
11 minute read
Ingolf Kanestrøm: Evolusjonens egne grenser


Evolusjonens egne grenser
Av Professor Ingolf Kanestrøm Universitetet i Oslo
Det er blitt etterlyst hvilke prosesser i evolusjonen som har ledet til kompliserte organer som for eksempel menneskets øye. Det vanlige svaret er at tilfeldige variasjoner (mutasjoner) med naturlig utvalg kan frembringe alle de organer og individer vi observerer i dag, om en bare har nok tid. Tiden er ikke den eneste kritiske faktor, men produktet av tid og antall individer. Dette er viktig, for det betyr at når vi studerer utviklingen av mikroorganismer, som visse arter bakterier, vil utviklingen over noen tiår kunne tilsvare utviklingen av mennesket i
Malariamygg Malariamygg Malariamygg Malariamygg Malariamygg
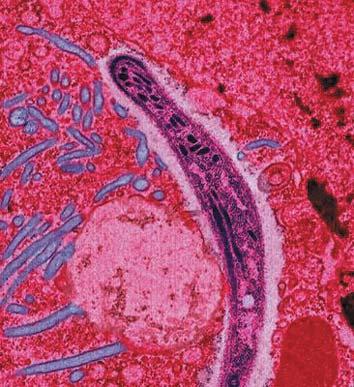
Malariaparasitt Malariaparasitt Malariaparasitt Malariaparasitt Malariaparasitt hele menneskets historie. Dette skulle gi oss en mulighet til å studere evolusjon i laboratorier over noen tiår.
Michael Behe, professor i biokjemi ved Lehigh University, Pensilvania, stiller spørsmål om riktigheten av evolusjonslærens grunnsetninger. For å finne svaret må utviklingslæren stilles på inngående prøve. Vil ideene om tilfeldig mutasjon, naturlig utvalg og felles avstamming bestå en kritisk prøve? Vi skal her se på de to førstnevnte og basere oss på boken The Edge of Evolution av Behe. Den kom ut i 2007.
Det mest kritiske elementet i utviklingslæren er tilfeldige variasjoner. I følge Darwin er den eneste måten en plante eller et dyr kan bli bedre tilpasset omgivelsene enn deres slektninger, er å opprettholde en fordelaktig mutasjon. Dersom mutasjonen gjør organismen sterkere, raskere etc, kan naturlig seleksjon overta og bidra til at antall avkom øker. Men inntil en vilkårlig mutasjon inntreffer, må naturlig seleksjon bare sitte og vente. Forskningsresultater gjennom det siste hundreåret viser at tilfeldige mutasjoner koplet med naturlig utvalg kan modifisere livet betydelig. Men den samme forskning viser at tilfeldige mutasjoner har en ekstremt begrenset virkning.
Malaria Malaria har virket som en svøpe over store deler av verden. Mange steder har malaria tatt livet av halvparten av barnepopulasjonen før de ble fem år gamle. Midt i forrige århundre kom det nye medisiner mot sykdommen, og håpet om at malaria kunne utryddes, ble tent. Men etter noen år utviklet malariaparasitten resistens mot medisinen. Nye medisiner ble utviklet, men uten varig effekt. Så i stedet for håpet om at menneskene kunne utrydde malaria, oppstod det frykt for at malaria kunne utrydde menneskene i deler av verden. Dette indikerer at utviklingen er ubarmhjertig, den avfeier virkningen av moderne medisin.
Genene til menneskene og malariaparasitten har ligget i strid i tusener av år. I denne kampen har enhver mutasjon som har gitt den ene en fordel overfor den andre, blitt favorisert av naturlig utvalg og fått økt avkom.
Malariaparasitten er et encellet individ som bæres av malariamyggen. Når et menneske blir bitt av myggen, går parasitten inn i blodsystemet og blir transportert til leveren. Der stopper parasitten og formerer seg. Så går den tilbake til blodstrømmen og angriper de røde blodlegemene. Denne prosessen fortsetter, og i løpet av noen få dager kan billioner av parasitter bli dannet, og de kan ødelegge en stor del av offerets blod.
Over århundre har menneskenes gener prøvd forskjellige forsvarsmetoder overfor malaria. Vitenskapen har lært en god del av denne forsvarskrigen, og noen resultater er uventet. Striden mellom parasittene og verten som av en rekke darwinister har blitt beskrevet som et produktivt rustningskappløp med forbedringer på begge sider, viser seg å være en skyttergravskrig der forholdene blir dårligere.
Sigdcelle karaktertrekk En grunnleggende prosess hos alle vesener er absorpsjon av oksygen fra omgivelsene, slik at cellene får tilført oksygen og stoffskiftet kan finne sted. Hos menneskene skjer denne prosessen i hemoglobinet (proteiner) i de røde blodlegemene.
Hemoglobin har fire kjeder av aminosyrer, hvorav to og to er like. Så hender det en sjelden gang at en aminosyre tar plassen til en annen (i posisjon nr. 6 av 146). Da får vi det vi kaller sigdcellehemoglobin. Dette gjør at hemoglobinmolekylet blir mindre oppløselig. Blodlegemene som normalt er skiveformet, blir sigdformet når de avgir oksygen. En konsekvens er at molekylene får en egenskap som ligner magneter. De hekter seg sammen, flere og flere. De koagulerer til en gelelignende masse i de røde blodcellene. Dermed kan sigdcellene blokkere de minste blodårene og tappe omkringliggende vev for oksygen. Folk som har arvet et slik gen fra bare den ene av foreldrene, sies å ha sigdcelle karaktertrekk. Det er ikke noe fatalt med denne arven. Men sett fra malariaparasittens ståsted har noe gått fullstendig galt. Etter som malariaparasitten forsyner seg av blodet, forandrer de røde blodcellene seg. I stedet for at de individuelle hemoglobincellene dytter (red.: puffer) på hverandre pga molekylær bevegelse, fester de seg til hverandre. Snart vil hele vesken av hemoglobinløsninger begynne å stivne. Spredningen av denne gelemassen vil presse mot parasitten og membranen av de røde blodcellene og forandre formen på disse. Når gelen følger blodstrømmen, og før parasitten får tid til å feste seg, vil den infiserte cellen nå milten. Milten griper tak i den infiserte cellen og ødelegger den og parasitten. Pasienten vil overleve angrepet takket være kampen som hemoglobinet har utkjempet. Ut fra darwinismen skulle man tro at alle mennesker der malaria herjer, skulle ha denne genetiske “feilen”. Men det er en hake knyttet til fenomenet. Sigdcelleanemi Dersom en arver sigdegenskapen fra begge foreldrene, kan en utvikle full anemi, og de fleste som barn. I Amerika bærer omlag 10 % av afroamerikanerne sigdegenskapene, og omlag 100 000 har anemi. Nå kan vi se hvorfor sigdcelleanemi har vart så lenge? Hvorfor er den ikke forsvunnet? Muta- Sigdceller Sigdceller Sigdceller Sigdceller Sigdceller sjonen har både positive og negative konsekvenser. Så lenge man arver genet fra bare den ene av foreldrene, er mutasjonen positiv fordi den beskytter mot malaria. Når en arver det fra begge foreldrene, er det dødelig. Utbredelsen av sigdcellehemoglobin vil derfor avhenge av en form for likevekt mellom de positive effektene og de dødelige konsekvensene.
Hemoglobin-C (HbC) Dette er betegnelsen på hemoglobin hvor det har skjedd en mutasjon. Også her er det bytte av aminosyrer i 6. posisjon i en av aminokjedene. Men i dette tilfelle er de to aminosyrene svert like unntatt den elektriske ladning som har motsatt fortegn. Det viser seg at HbC-hemoglobin ikke størkner på samme måte som sigdhemoglobin. Det hjelper mennesker å bekjempe malaria, men årsaken er ikke helt forstått. For å få full effekt mot malaria, er det nødvendig med to kopier av C-genet. HbC-hemoglobin forårsaker ikke så store problemer som sigd-hemoglobin. HbC-hemoglobinet er begrenset til noen få regioner i Afrika.
Både sigdcelle- og HbC-mutasjoner er i hovedsak skadelige mutasjoner da de reduserer menneskekroppens funksjoner. Begge fremkaller anemi og andre ugunstige effekter. Under gunstige forhold ville de ikke få fotfeste i en menneskelig populasjon. Men i den desperate situasjon der for eksempel malaria truer menneskene, kan det være gunstig å akseptere noen ulemper for å holde “fienden” ute.
Mutasjonene samarbeider ikke for å bygge et mer komplekst vekselvirkende biokjemisk system. Sigd-mutasjonen og HbC-mutasjonen er gjensidig utelukkende siden de finner sted i samme posisjon i aminokjeden. De kan således ikke gjøre noe sammen. Ingen av mutasjonene finner sted i immunsystemet, systemet som normalt er ansvarlig for å bekjempe mikroskopiske inntrengere. De to mutasjonene virker ikke koherent, samspillet mellom de muterte og normale genene er kaotisk. Sigdcellene spres raskt og etabler seg som “kongen på haugen” før HbC-cellene kan starte. Bærere av sigdceller vil være best stilt ved å gifte seg med noen som ikke er bærere, mens bærere av HbC vil være best tjent med å gifte seg med en slektning som og har HbC. Det kaotiske samspillet av gener er ikke konstruktivt. Erfaringer viser at mange løse tråder blir tvunnet sammen
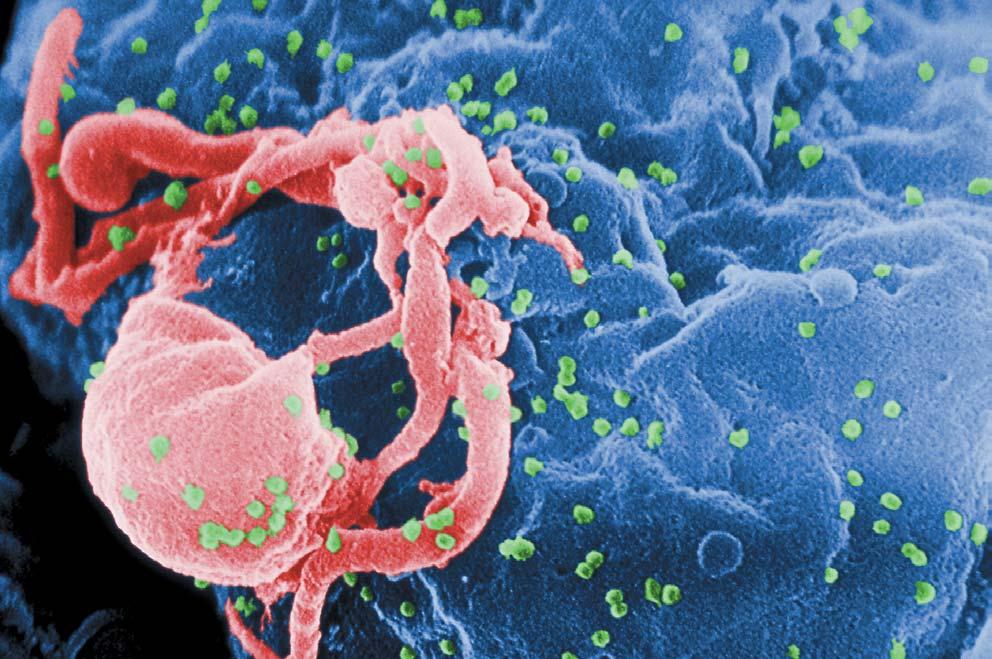
Viruset HIV Viruset HIV Viruset HIV Viruset HIV Viruset HIV-1 (grønne punkter) i aksjon. F -1 (grønne punkter) i aksjon. F -1 (grønne punkter) i aksjon. F -1 (grønne punkter) i aksjon. F -1 (grønne punkter) i aksjon. Fargene på bildet er kunstige. (F argene på bildet er kunstige. (F argene på bildet er kunstige. (F argene på bildet er kunstige. (F argene på bildet er kunstige. (Fra Wikipedia.) ra Wikipedia.) ra Wikipedia.) ra Wikipedia.) ra Wikipedia.)
og resulterer i en gordisk knute. Spørsmålet blir da: er det dit den darwinistiske evolusjon skal føre oss?
HIV og evolusjon Viktig informasjon kan hentes fra studier av HIV-virus som fremkaller AIDS. Mengden av genetisk informasjon i HIV-viruset er mindre enn en 1000 del av informasjon i malariaparasitten. Mutasjoner i virus foregår om lag 10 000 ganger raskere enn i celler. Derfor vil i middel en kopi av viruset inneholde en mutasjon fra dets foreldre. Derfor kan darwinistisk utvikling neppe gå fortere enn i HIV-viruset.
En antar at en person infisert av HIV bærer mellom en og ti milliarder virus. Generasjonstiden for virus er en dag eller to. Over en 10-års periode vil en HIV-pasient produsere mer enn 1 000 generasjoner HIV, eller opp til ti billioner virus. Da en antar at det er femti millioner HIV-smittede i verden, kan en anta at det er blitt produsert 1020 (1 med 20 nuller bak) virus. Det store antall kopier av HIV-viruset må derfor antas å inneholde tilnærmet alle tenkelige mutasjoner. Men hva har utviklingen forbundet med alle disse mutasjonene gjort. Svaret må bli: Svært lite!
AIDS har tatt livet av millioner av mennesker, parert menneskets immunsystem og utviklet resistens mot medisin. Med få unntak entrer HIV-viruset immunsystemet ved å binde seg sterkt til et av de mange proteinene i deres overflate for så å strekke seg for å binde seg til et annet protein. Denne prosessen fortsetter, og hundrede av milliarder muterte virus gjør det fortsatt på samme måte. Dersom et mutert virus hadde utviklet evner til å innta andre slag celler ved å binde seg til andre proteiner, kunne det formere seg mer effektivt og utkonkurrere sine søsken. Men det har ikke skjedd. Heller har det ikke skjedd mye på molekylnivå. Ingen fancy triks som opptrer i darwinistisk spekulasjoner, har blitt til noen spesiell nytte for HIV.
Hva med hurtig utvikling av resistens mot medisin? Viser dette at darwinistisk evolusjon er effektiv? Forskning viser at HIV benytter det samme moderate triks som maleria benytter, en punktmutasjon for å redusere bindingen av giften til det sykdomsfremkallende målet. Det kan være en forandring av bare en aminosyre i posisjon nr. 184 i et spesielt HIV-enzym som danner en liten dump som vekselvirker med en medisin.
E.coli-bakterier Studier av maleria og HIV utgjør gode muligheter for å fremskaffe evidens for hva evolusjonen kan utrette. Årsaken er det store antall. Dess større antall organismer som finnes, dess større er sjansen for at en positiv mutasjon kan opptre og bli fanget opp av naturlig selek-
E E E E E.coli-bakterier .coli-bakterier .coli-bakterier .coli-bakterier .coli-bakterier
sjon. Men det finnes tilfeller der mulig evolusjon kan studeres i laboratorier. Ved Michigan University startet et laboratorieforsøk for mer enn ti år siden. E.coli-bakterier er en favorittorganisme når det gjelder laboratoriestudier. Bakteriene er lette å dyrke, og de har en kort generasjonstid, ca 20 minutter under gunstige forhold. Hele arvematerialet til bakterien er kartlagt.
Til forskjell fra maleria og HIV som må klare seg selv i naturen og slåss mot det menneskelige immunforsvar, blir E.coli degget med (da.: opflasket) i laboratoriet. De har stabile omgivelser, daglig føde og ingen fiender. Noen vil hevde at stabile omgivelser ikke vil stimulere til evolusjon. Men en av de viktigste faktorene i organismens omgivelser er tilstedeværelsen av andre organismer. Selv i kontrollert laboratoriekultur der bakteriene er varme og velfødde, vil individer som formerer seg hurtigst eller utkonkurrerer de andre, dominere populasjonen. I dette tilfellet ble E.coli dyrket i flasker som nådde sin kapasitet etter seks-sju doblinger. Hver dag ble en del av bakteriene overført til en ny flaske. Nå har 30 000 generasjoner av E.coli, ekvivalent med en million år i menneskets historie, blitt fødd og har dødd i laboratoriet. I hver flaske hadde populasjonen vokst til omkring 500 millioner, totalt i eksperimentet til ca. ti billioner bakterier. Men analyser viser at ikke noe fun-
damentalt nytt er blitt dannet, ingen ny protein-protein vekselvirkning, ingen ny molekylær maskin. Noen populasjoner av bakterier har mistet evnen til å reparere DNA. En av de mest nyttige mutasjonene var bakterienes tap av evnen til å lage sukker av ribose. Etter en tid rant gunstige forandringer fra eksperimentet ut i sand.
Malariaparasitter, HIV og E.coli er svært forskjellig fra hverandre. De strekker seg fra det enkle til det komplekse, de har forskjellig livssyklus, og de dekker tre forskjellige livsdomener, eukaryot (individer med celler som har kjerner), virus og prokaryot (individer med celler uten kjerne). Alle tre forteller den samme historien om darwinistisk utvikling. Noen enkle forandringer i gammelt cellulare maskineri som kan være nyttig i vanskelige situasjoner, opptrer. De funger når det gjelder antibiotika resistens. De kan brenne “broer” når det kommer en hær av inntrengere. Men når det kreves flere sprang for at det skal ha en nyttig effekt, begynner vilkårlige mutasjoner å puste tungt. Å utøve enda flere sprang synes å være utenfor evolusjonens rekkevidde. Behes analyser av studier av malariaparasitter, HIV-virus og E.coli-bakterier viser at darwinistisk evolusjon kan bryte ned enkle molekyler, men den kan ikke skape noe fundamentalt nytt. Den kan ikke bygge opp kompliserte molekylære maskineri.





