(Ebook) Insect-Fungal Associations: Ecology and Evolution by Fernando E. Vega, Meredith Blackwell ISBN 9780195166521, 9781423785156, 0195166523, 1423785150
(Ebook) Cambridge IGCSE and O Level History Workbook 2C - Depth Study: the United States, 1919-41 2nd Edition by Benjamin Harrison ISBN 9781398375147, 9781398375048, 1398375144, 1398375047
A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research
Series Editors: Dee Carter · Anuradha Chowdhary
Joseph Heitman · Ulrich Kück
Yen-Ping Hsueh
Meredith Blackwell Editors
Fungal Associations
Third Edition
The Mycota
AComprehensiveTreatiseonFungias
ExperimentalSystemsforBasicandApplied Research
Volume9
Series Editors
Dee Carter, School of Life and Environmental Sciences, The University of Sydney, Sydney, NSW, Australia
Anuradha Chowdhary, Department of Medical Mycology, VP Chest Institute, University of Delhi, Delhi, India
Joseph Heitman, Duke University Medical Center, Durham, NC, USA
Ulrich Kück, General and Molecular Botany, Ruhr-University Bochum, Bochum, Germany
The fungi represent a heterogenous assemblage of eukaryotic microorganisms and have become favored organisms for research at the cellular and molecular level. Such research involvement has been stimulated by interest in the biotechnological application of fungi in processes related to industry, agriculture and ecology. Considering both yeasts and mycelial fungi, THE MYCOTA highlights developments in both basic and applied research and presents an overview of fungal systematics and cell structure. Foremost authorities in research on mycology have been assembled to edit and contribute to the volumes.
Yen-PingHsueh •
MeredithBlackwell Editors
FungalAssociations
ThirdEdition
Editors
Yen-Ping Hsueh
Institute of Molecular Biology, Academia Sinica
Taipei, Taiwan
Meredith Blackwell
Department of Biological Sciences
Louisiana State University Baton Rouge, LA, USA
Department of Biological Sciences
University of South Carolina Columbia, SC, USA
ISSN 2945-8048ISSN 2945-8056 (electronic)
The Mycota
ISBN 978-3-031-41647-7ISBN 978-3-031-41648-4 (eBook) https://doi.org/10.1007/978-3-031-41648-4
# The Editor(s) (if applicable) and The Author(s), under exlusive licence to Springer Nature Switzerland AG 2001, 2012, 2024
This work is subject to copyright. All rights are solely and exclusively licensed by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. The publisher, the authors, and the editors are safe to assume that the advice and information in this book are believed to be true and accurate at the date of publication. Neither the publisher nor the authors or the editors give a warranty, expressed or implied, with respect to the material contained herein or for any errors or omissions that may have been made. The publisher remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This Springer imprint is published by the registered company Springer Nature Switzerland AG The registered company address is: Gewerbestrasse 11, 6330 Cham, Switzerland
Paper in this product is recyclable.
Series Preface
The Mycota is an encyclopedic book series published by Springer as a comprehensive treatise spanning the Fungal Kingdom through a focus on fungi as experimental systems for basic and applied research. Articles in this series present introductory information in a comprehensive manner to broaden our general knowledge, along with state-of-the-art information on selected topics in fungal biology. Thus, these articles serve as a timeless reference source for historically relevant discoveries in fungal biology to drive further advances in the field.
The Mycota was founded in 1994 by Karl Esser and Paul Lemke. Since then 15 volumes have been published, with several now in their third edition. The steadily growing interest in The Mycota,reflected in particular by the high number of e-book downloads, encouraged Springer to continue this publication with a new international Series Editorial Board beginning in 2022.
Historically, the study of fungi originated as a sub-discipline of botany and was a largely descriptive discipline until the early nineteenth century. Subsequent experimental research has propelled the field of mycology, and achievements in the genetics and molecular biology of fungi have benefited studies in the related fields of fungal biochemistry and biophysics, plant pathology, medical mycology, and systematics. Ground-breaking research has been carried out using fungi as recognized by the receipt of nine Nobel Prizes, including the revolutionary 1945 Nobel Prize in Physiology or Medicine for the discovery of penicillin and the foundational 2006 Nobel Prize in Chemistry for establishing the molecular basis of eukaryotic transcription. All such studies expand our knowledge of fungi and of fungal processes and improve our ability to understand, utilize, and control fungi for the benefitof humankind.
Fungi have invaded every conceivable ecological niche. Saprobic forms abound, especially in the decay of organic debris. Pathogenic forms exist with both plant and animal hosts. Fungi even grow on other fungi. They are found in aquatic as well as soil environments, and their spores have been found in the air of extreme environments. Fungi can be variously associated with plants as symbionts in the form of lichens, mycorrhizae, and endophytes and also occur as overt pathogens. Association with animal systems varies; examples include the predaceous fungi that trap nematodes, the micro-fungi that grow in the anaerobic environment of the rumen, and medically important pathogens afflicting humans. Currently, taxonomists have identified about 150,000
species, but it is thought that over 90% of fungal species remain undescribed, with conservative estimates ranging from 2 to 5 million fungal species on earth.
From this perspective, there are new topics in fungal biology to explore, which will expand the current volumes and take them in new directions. Further, there will be new volumes in areas (e.g., Cryptomycota) that were not covered before.
Our understanding of the evolution of fungi is still incomplete and mainly based on species that can be grown in culture. But recent environmental DNA analyses have revealed a highly diverse form of eukaryotic life that branches with the Fungi, and thus the resulting and highly diverse clades were named the cyptomycota. The discovery of novel intermediate forms will redefine the fungal tree of life. Other topics will consider the adaptation of fungi to climate changes, the occurrence of fungal pathogens in the environment, and the dispersal of fungi during global pandemics.
Fungal constituents of the microbiome have received much less attention thus far, yet recent findings from clinical and animal studies clearly establish fungi as a significant component of the oral, gastrointestinal, pulmonary, and skin microbiomes. Finally, the relevance of fungal biology to society is reflected by the increasing number of fungal related human diseases.
In the history of pharmacology, fungi have always been sources of useful molecules for humans, but the diversity of molecules that can be obtained from fungi is still largely under explored. However, efforts in fungal genomics provide resources for data-driven genome mining and large-scale comparisons to explore the molecular repertoires produced by fungi. The result will be new compounds with applications in the pharmaceutical and agricultural science industries.
Fungi can also help the entire planet, and may for example be relevant in speci fic sectors, such as that linked to pollution from plastic. Fungi produce a wide range of enzymes that have the potential to break down the chemical bonds of plastic polymers, and in this context, the potential role of marine fungi in plastic degradation may be of major relevance. Finally, new biomaterials from fungal species may open the door to alternatives to fossilbased materials, and thus reduce environmental pollution.
For consistency throughout this series of volumes, the names adopted for major groups of fungi should be followed according to the following paper, which gives an overview of all of the orders in the fungal kingdom: https:// pubmed.ncbi.nlm.nih.gov/32660385/
We are grateful to Springer for continuing The Mycota series and are especially thankful to all the volume editors in selecting topics and assembling experts from diverse fields of fungal biology.
Sydney, AustraliaDee Carter Delhi, IndiaAnuradha Chowdhary Durham, USAJoseph Heitman Bochum, GermanyUlrich Kück July 15th, 2022
Volume Preface to the Third Edition
Fungi are heterotrophic eukaryotes with chitinous cell walls; some species are single-celled flagellates, yeasts, or yeast-like cells, and many species have complex structures composed of filaments known as hyphae. Except for spore-bearing mushrooms and other large reproductive structures, mycology’s charismatic megamycota (Blackwell 2011), most fungi go unseen except for their traces of deterioration and disease. Other characteristics of fungi include osmotropic nutrition whereby enzymes exported to the substrate dissolve organic nutrients that then are absorbed into the organism. Fungi also produce vitamins, sterols, and aromas, often important in their interactions and communication with other organisms. Fungi are an extraordinarily diverse group, in part because of myriad associations (Vega and Biedermann 2019) with almost all kinds of organisms including other fungi.
About a billion years ago populations of ancestral eukaryotic single-celled organisms diverged into fungi and animals, two dominant megadiverse groups we recognize today. Together with plants, fungi and animals constitute the “fauna, flora, and funga,” a triumvirate of the most prominent organisms on Earth (Pfister 2020; Berbee and Blackwell 2023). Fungal associations range widely from simple co-occurrence, perhaps the result of only accidental spore dispersal, to complex multipartite symbioses in the broad sense, including mutualistic, commensalistic, and parasitic lifestyles.
This volume looks at the results of a billion years of fungal associations revealed by current methods:
• Many animals and plants are protected from fungi by preadapted and evolved defenses.
• Some fungi have been able to have such intimate contact with other organisms that their DNA has been exchanged.
• The dogma that obligately symbiotic fungi cannot be grown in axenic culture is being investigated with the help of genomes.
• Aromatic volatile compounds lure or repel other organisms in what would seem to be a confusing mélange of bacterial, animal, and plant compounds in the environment.
• Examples of mycoviruses and their influence on fungal phenotypes are remarkable; additional associations and their effects await discovery. [Meredith Blackwell was especially interested to see a reference to virus-
infected chestnut trees in Michigan, perhaps those discovered by an undergraduate at Hope College, Holland, Michigan, where she once taught.]
• Classic textbook examples of fungal associations have been completely updated by the application of traditional and new molecular techniques. These chapters cover lichens, including a model lichen, arbuscular mycorrhizal fungi, ectomycorrizal fungi, and an endophyte rendered much less toxic.
• Also included are fungal predators of nematodes, fungi that control the minds of insects, and even more fungal killers of insects.
• The volume closes with a pharmacopeia of secondary metabolites synthesized by sclerotium-producing Xylarias in termite galleries — metabolites with pharmaceutical potential.
Development of new technologies precedes advancements in our science. Evidence of progress brought about by the polymerase chain reaction (PCR), especially applied to fungi (White et al. 1990), and all the resulting improvements in molecular biology are evident in the research discussed in this volume. As we completed editing these chapters, arti ficial intelligence (AI) crept up and suddenly burst forth into the scienti fic literature and mass media, even into the hearing rooms and statutes of governemental bodies. The study of fungal associations and related fields is poised to adopt the use of AI through today’s increased access to supercomputers, ever-expanding databases (including the proliferation of scientific literature), and a need to recognize complex patterns; perhaps AI even engenders a willingness to have complicated decisions made for us and eases the demands to undertake repetitive tasks. We anticipate that as successive technologies are developed and improved, future studies of fungi and the organisms with which they associate will continue to reap benefits.
We appreciate the help of our colleagues Donald H. Pfister, Joseph W. Spatafora, and Marc Rapport for editorial assistance. We are especially grateful to our contacts at Springer Nature, Sabine Schwarz, Executive Editor, Life Sciences & Biomedicine Books, and Bibhuti Bhusan Sharma, Project Coordinator (Books), for their unlimited support from beginning to end during the preparation of Mycota 9: Fungal Associations. Without them two neophytes would not have managed. Our helpful and patient families are of course gratefully acknowledged.
Columbia, SCMeredith Blackwell Taipei, TaiwanYen-Ping Hsueh 11 June 2023
Literature
Blackwell M (2011) The Fungi: 1, 2, 3, 5.1 million species? Biodiversity special issue. Am J Bot 98:426–438. doi: 10.3732/ajb.1000298
Berbee M, Blackwell M (2023) The origin of fungi: A billion years of functional and morphological diversification. In: Harper D, Seberg O (eds) The origins of all things. Munksgaard, Copenhagen, pp 231–277
P
fister DR (2020) Fantastic Funga: why language matters. Deep Funga Blog. https://fundis.org/resources/blog/127-fantastic-funga-why-languagematters
Vega FE, Biedermann PH (2020) On interactions, associations, mycetangia, mutualists and symbioses in insect-fungus symbioses. Fungal Ecol 44: 100909. doi:10.1016/j.funeco.2019.100909
White TJ, Bruns TD, Lee SB, Taylor JW (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic, New York
1 When Plants and Animals First Met Fungi: Insights from the Evolution of Host Immune Systems ... .... ... .. . 1
Emily D. Trudeau and Mary L. Berbee
2 Metabolic Constraints and Dependencies Between “Uncultivable” Fungi and Their Hosts 33
Saleh Rahimlou, C. Alisha Quandt, and Timothy Y. James
3 Horizontal Gene Transfer in Fungi and Its Ecological Importance 59
Carla Gonçalves, Chris Todd Hittinger, and Antonis Rokas
4 An Overview of Fungal Volatile Organic Compounds (VOCs) 83 Samantha Lee, Richard Hung, and Joan W. Bennett
5 Viruses that Affect Phenotype and Fitness of Fungi 113 Bradley I. Hillman and Massimo Turina
6 Lichens 145 Martin Grube
7 Umbilicaria muhlenbergii: A Model for Studying Molecular Mechanisms Regulating Initial Fungal Symbiotic Interactions with Algal Cells ....... ....... .. . 181 Yanyan Wang, Xinli Wei, Yuting Hu, and Jin-Rong Xu
8 After Air, Light, and Water, the Next Most Important Thing Is Grass: An Introduction to the Epichloë–Grass Symbiosis 195
Daniel A. Bastías, Emma R. Applegate, Pedro E. Gundel, Linda J. Johnson, Wade J. Mace, Christina D. Moon, Sushma Prakash, and Stuart D. Card
9 Signals and Host Cell Remodeling in Arbuscular Mycorrhizal Symbiosis 231 Andrea Genre, Serena Capitanio, and Paola Bonfante
10 Masters of Manipulation: How Our Molecular Understanding of Model Symbiotic Fungi and Their Hosts Is Changing the Face of “Mutualism” 249
Jonathan M. Plett, Annegret Kohler, and Francis Martin
11 Nematode-Trapping Fungi and Caenorhabditis elegans as a Model System for Predator–Prey Interactions 273
Guillermo Vidal-Diez de Ulzurrun, Sheng-Chian Juan, Tzu-Hsiang Lin, and Yen-Ping Hsueh
12 When a Mind Is Not Its Own: Mechanisms of Host Behavior Control by Parasitic Fungi ..
Carolyn Elya
13 Genetics and Infection Biology of the Entomopathogenic Fungi 309
Song Hong, Junmei Shang, Yanlei Sun, and Chengshu Wang
14 Xylaria Sclerotia Formed Within Termite Nests: A Review of Their Biology and Human Uses 333 Huei-Mei Hsieh and Yu-Ming Ju
Editors and Contributors
About the Series Editors
Dee Carter is Professor of Microbiology at the University of Sydney. She has a degree in microbiology and biochemistry from Otago University, New Zealand, and a PhD from Imperial College, London, UK. After postdoctoral fellowships in Montpellier, France, and UC Berkeley, California, USA, she moved to Australia in 1995 to take up a lectureship at the University of Sydney. Dee’s research has encompassed (1) the population genetics and ecology of medically important fungi, including yeast and mold pathogens; (2) responses of fungi to host and antifungal stress, including the production of variant morphological forms; and (3) the transcriptome and proteome response to antifungal therapy, particularly during synergistic interactions between antifungals and natural products. As well as research, Dee teaches microbiology including mycology, epidemiology, and molecular biology at Sydney University.
Anuradha Chowdhary is Professor of Medical Mycology at the Vallabhbhai Patel Chest Institute, University of Delhi, India. She received her medical degree in 1992 and MD (Microbiology) degree in 1996 from the University of Delhi, India, and PhD from the Faculty of Medical Sciences, Radboud University Medical Center, Netherlands. Dr Chowdhary’s research interests include molecular ecology and population genetics of pathogenic fungi, antifungal drug resistance
mechanisms, especially Aspergillus and Candida spp., and the epidemiology of systemic mycoses. She is currently working on molecular epidemiology of Candida auris, terbinafine-resistant dermatophytes, and azole-resistant Aspergillus fumigatus.
Joseph Heitman is James B. Duke Professor and Chair, Department of Molecular Genetics and Microbiology, Duke University, Durham, NC, USA. His research studies model and pathogenic fungi addressing fundamental questions of scientific and medical importance. Pioneering studies with Baker ’s yeast revealed how immunosuppressive natural products interdict signaling cascades via FKBP12-drug complexes, and discovered TOR as a globally conserved nutrient sensor targeted by the immunosuppressive, antiproliferative drug rapamycin. His research discovered unisexual reproduction of pathogenic microbes, with implications for pathogen emergence, how sex generates diversity, and how sex evolved. Dr Heitman’s lab has further developed genetic and genomic approaches elucidating molecular principles of fungal virulence, identifying therapeutic targets, and illustrating convergent evolution of fungal mating-type loci with mammalian, insect, and plant sex chromosomes, defined the calcium-activated protein phosphatase calcineurin as a globally conserved fungal virulence factor, and elucidated functions of RNAi in microbial pathogen genome integrity, hypervirulent outbreak lineages, and drug resistance via epimutation.
Ulrich Kück is Professor of General and Molecular Botany at the Ruhr-University in Bochum, Germany. He graduated in Biology and Chemistry, and has a long-standing experience in the molecular biology of fungi, algae, and plants. His research with fungi has focused on two general aspects: first, genetic engineering of the secondary metabolism of biotechnically relevant filamentous fungi, including functional
genomics and proteomics, and second, molecular genetic analysis of cellular growth in filamentous fungus, with a focus on sexual development. In particular, he is interested in the function of mating-type loci, and the involvement of conserved signaling complexes, such as the striatin-interacting phosphatases and kinases (STRIPAK) complex, in the control of cellular and developmental processes.
About the Volume Editors
Yen-Ping Hsueh was born in Taipei, Taiwan. Biology has always been her favorite subject in school so she followed her interest and majored in Plant Pathology and Microbiology at the National Taiwan University as an undergraduate. She received BSc (2001) and MSc (2003) degrees, and moved to Duke University to pursue her PhD under the guidance of Dr Joseph Heitman. Her PhD work mainly focused on the sexual cycle of the human fungal pathogen Cryptococcus neoformans. After receiving her PhD in Molecular Genetics and Microbiology from Duke University in 2008, she continued to work with Heitman briefly and then moved to Caltech to start her postdoctoral training with Dr Paul W. Sternberg in 2010. Her postdoctoral work was an investigation of the interactions between nematode-trapping fungi and the model nematode C. elegans.
In 2015, Hsueh returned to Taiwan, to begin her independent career at the Institute of Molecular Biology in Academia Sinica where she is currently Associate Research Fellow. Her laboratory studies the predator–prey interactions between carnivorous fungi and the model nematode Caenorhabditis elegans. Members of the Hsueh lab explore different aspects of the biology of these predator–prey systems and are striving to understand the mechanisms that underlie their interactions and coevolution. They hope to harness the nematocidal potential of the carnivorous
fungi, which will facilitate the discovery of a novel treatment or biocontrol method to combat parasitic nematode infections in plants, animals, and humans in the future. Because of her original research, Hsueh has been recognized as an EMBO Global Investigator, EMBO Young Investigator, and more recently, a Taiwan Outstanding Women in Science Young Talent Award and is an active participant in the international scientific community.
Meredith Blackwell born in Abbeville, Louisiana, spent her early years on the banks of the Vermilion Bayou and attended public schools in Louisiana. As an undergraduate pursuing liberal arts interests, a deep-sea fishing trip led her to become a biology major (BS, University of Louisiana, 1961). She studied ichthyology at the University of Alabama (MS, 1963) and began a PhD at the University of Texas at Austin in 1964. At Texas, she eventually came under the guidance of C. J. Alexopoulos, and her enduring interest in Mycology developed (PhD, 1973). During postdoctoral research with Henry Aldrich and James Kimbrough at the University of Florida, she was introduced to insect-associated fungi. She spent the first 6 years of her career primarily in a teaching position at Hope College, Holland, Michigan. The next 33 years (1981–2014) were spent in the Department of Biological Sciences, Louisiana State University, where she attained the rank of Boyd Professor. Her research supported by NSF included studies of desert myxomycetes, phylogeny of Laboulbeniomycetes and other insect ectoparasites, and the taxonomic and geographic distribution of insect-associated yeasts. Since 2014, she has lived in Columbia, South Carolina, where she is an affiliate in the Department of Biological Sciences at the University of South Carolina. Blackwell is a former president of the Mycological Society of America (MSA) and the International Mycological Association (IMA). She is a fellow of the British Mycological Association, MSA, IMA, and the American Academy of Arts and Sciences. She has received several mycological awards and is proud to share her name with a number of fungal taxa.
Contributors
Emma Applegate Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Daniel Bastias Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Joan W. Bennett Department of Plant Biology, Rutgers, The State University of New Jersey, New Brunswick, NJ, USA
Mary Berbee Department of Botany, University of British Columbia, Vancouver, BC, Canada
Paola Bonfante Dipartimento di Scienze della Vita e Biologia dei Sistemi, Università di Torino, Torino, Italy
Stuart Card Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Serena Capitanio Dipartimento di Scienze della Vita e Biologia dei Sistemi, Università di Torino, Torino, Italy
Carolyn Elya Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA, USA
Andrea Genre Dipartimento di Scienze della Vita e Biologia dei Sistemi, Università di Torino, Torino, Italy
Carla Gonçalves Department of Biological Sciences and Evolutionary Studies Initiative, Vanderbilt University, Nashville, TN, USA
Martin Grube Institute of Biology, University of Graz, Graz, Austria
Pedro Gundel Instituto de Ciencias Biológicas, Talca University, Talca, Chile
Bradley Hillman Plant Biology, Rutgers University, New Brunswick, NJ, USA
Chris Todd Hittinger Laboratory of Genetics, DOE Great Lakes Bioenergy Research Center, Center for Genomic Science Innovation, Wisconsin Energy Institute, J.F. Crow Institute for the Study of Evolution, University of Wisconsin-Madison, WI, USA
Song Hong Laboratory of Insect Developmental and Evolutionary Biology, CAS Center for Excellence in Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai, China
Huei-Mei Hsieh Institute of Plant and Microbial Biology, Academia Sinica, Nankang, Taipei, Taiwan
Yen-Ping Hsueh Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan
Richard Hung Mycorrhizal Applications, St. Louis, MO, USA
Timothy Y. James Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI, USA
Linda Johnson Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Yu-Ming Ju Institute of Plant and Microbial Biology, Academia Sinica, Nankang, Taipei, Taiwan
Sheng-Chian Juan Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan
Annegret Kohler UMR 2UMR INRA/UHP 1136, Interactions Arbres/ Micro-organismes, Centre INRA de Nancy, Champenoux, France
Samantha Y. Lee Graduate Women in Science, Mullica Hill, NJ, USA
Tzu-Hsiang Lin Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan
Wade Mace Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Francis Martin 2UMR INRA/UHP 1136, Interactions Arbres/Microorganismes, Centre INRA de Nancy, Champenoux, France
Christina Moon Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Jonathan M. Plett Hawkesbury Institute for the Environment, Western Sydney University, Richmond, NSW, Australia
Sushma Prakash Resilient Agriculture, AgResearch Limited, Grasslands Research Centre, Palmerston North, New Zealand
Alisha Quandt Department of Ecology and Evolutionary Biology, University of Colorado, Boulder, CO, USA
Saleh Rahimlou Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI, USA
Antonis Rokas Department of Biological Sciences and Evolutionary Studies Initiative, Vanderbilt University, Nashville, TN, USA
Junmei Shang Laboratory of Insect Developmental and Evolutionary Biology, CAS Center for Excellence in Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai, China
Yanlei Sun Laboratory of Insect Developmental and Evolutionary Biology, CAS Center for Excellence in Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai, China
Emily Trudeau Department of Botany, University of British Columbia, Vancouver, BC, Canada
Massimo Turina IPSP-CNR, Torino, Italy
Guillermo Vidal-Diez de Ulzurrun Institute of Molecular Biology, Academia Sinica, Nankang, Taipei, Taiwan
Chengshu Wang Key Laboratory of Insect Developmental and Evolutionary Biology, CAS Center for Excellence in Molecular Plant Sciences, Shanghai Institute of Plant Physiology and Ecology, Chinese Academy of Sciences, Shanghai, China
School of Life Science and Technology, ShanghaiTech University, Shanghai, China
Yan-Yan Wang State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences, Beijing, China
Jin-Rong Xu Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN, USA
When Plants and Animals First Met Fungi: Insights from the Evolution of Host Immune Systems
EmilyD.TrudeauandMaryL.Berbee
Abstract
Fungi interact with plants and animals, leading variously to mutually beneficial partnerships or to devastating disease. Reflecting the importance of plant–fungal and animal–fungal interactions, the innate immune systems of terrestrial plants and animals detect fungi using inherited recognition receptors. Upon recognition, the receptor proteins stimulate signaling pathways that foster intimate mutualisms or trigger robust, multi-level innate defenses against parasitism. To provide points of discussion about how the evolutionary history of host recognition may have unfolded, we analyze the phylogenetic distributions of pattern-recognition receptors important in plant and animal immunity, and of two groups of enzymes important in defenses: chitinases in plants and β-glucanases in animals. Our analysis takes advantage of recently sequenced genomes of early-diverging host plants and animals. We draw upon experimental studies using model organisms that reveal how some host plants and animals perceive fungi. To infer functional homology, we explore recent studies using experimental approaches to analyze binding
E. D. Trudeau · M. L. Berbee (✉)
Department of Botany, University of British Columbia, Vancouver, BC, Canada
interactions between host proteins and pathogen molecules. We contend that phylogenetic ages of the host receptors provide a window, imperfect but intriguing, into the origins of the host systems that have been detecting fungi over hundreds of millions of years of geological time.
Reconstructing evolutionary histories of plant and animal proteins involved in sensing and responding to fungi provides a genomic basis upon which to speculate about how and when fungi first interacted with plants and animals. When fungi meet plant or animal hosts, myriad complex mechanisms mediate interactions. Host receptors recognize fungal molecules, setting in motion complex signaling cascades to mount a defense or to initiate mutualistic symbiosis.
The mechanisms underlying fungus–plant or fungus–animal interactions have important implications, both positive and negative. Fungal
# The Author(s), under exclusive license to Springer Nature Switzerland AG 2024 Y.-P. Hsueh, M. Blackwell (eds.), Fungal Associations, The Mycota 9, https://doi.org/10.1007/978-3-031-41648-4_1
1
pathogens devastate over 125 million metric tons of major crop plants including rice, wheat, and maize every year and account for over 25% of crop losses worldwide (Boddy 2016). Arbuscular mycorrhizal symbiosis, thought to have evolved concurrently with the first land plants, likely had a role in helping early plants to become established on land, thereby shaping the first terrestrial ecosystems (Delaux and Schornack 2021).
Fungi in humans range from causative agents of life-threatening, systemic diseases to commensals, such as Candida spp., which can be asymptomatic colonizers of mucous membranes. Some mammals, such as ruminants and other herbivores, derive benefits from a mutualistic relationship with gut fungi. These fungi possess the remarkable ability to break down plant cellulose, which is typically indigestible, into digestible sugars (Gordon and Phillips 1998). Mammals and birds are protected from most potentially deadly fungal pathogens by body temperatures above 35 °C, by slightly alkaline body fluids (most fungi prefer slightly acidic substrates), and by skin surfaces that resist fungal attachment (Casadevall 2005). However, selection for additional layers of antifungal protection must have exerted continuing force through animal evolution. This is evident from medical records and laboratory experiments showing that pathogenic fungi rage out of control when our layers of immune protection are compromised (Brown et al. 2018). Especially in parts of the world where availability of sophisticated antiviral drugs is limited, fungal infections kill 500,000 HIV/AIDS patients per year (Limper et al. 2017) because virus-induced faults in adaptive immunity reduce antifungal protection.
Immune systems vary across groups of organisms. Adaptive immunity is widely shared by vertebrates and appears to have originated among early jawed fishes (Buchmann 2014; Rombout et al. 2005). Adaptive immunity results when an individual’s exposure to an antigen creates a “molecular memory,” which can take the form of antibodies speci fic to the antigen. Vaccination or exposure to infection triggers adaptive immunity. The molecular memory may
last a few months or a lifetime, but it is not passed to the next generation.
Innate immunity, the subject of this chapter, is based on genetically inherited receptors that recognize and bind ligands, which are molecules characteristic of various microbes. Distinguishing among molecules from interacting microbes enables eukaryotic hosts to initiate intracellular signaling, leading to partnerships with mutualists, tolerance of commensals, or the detection and destruction of pathogens. In vertebrates, response to microbes involves coordination between innate and adaptive immune systems. However, the vast majority of eukaryotes including plants and invertebrates have only the more ancient innate immune systems.
We focus on a first line of innate defense that starts when host proteins, pattern-recognition receptors at the cell surface, detect microbeassociated molecular patterns (Sanchez-Vallet et al. 2014; Hatinguais et al. 2020). The hosts’ receptors typically target microbial cell coverings. Their ability to distinguish among microbes usually involves their binding preferences. Cell coverings chitin, mannose, and β-1,3-1,6-glucan in fungi (Gow et al. 2017); lipopolysaccharides and peptidoglycans in Gramnegative bacteria; and peptidoglycans and teichoic acid in Gram-positive bacteria (Fisher and Mobashery 2020) are ancient molecules, but their organization and components vary and overlap, due in part to continuing selective pressure from pathogens and symbionts. Here, we focus on fungal recognition, but the same or similar host receptors may also detect oomycetes, nematodes, and other organisms that share similar cell surface biochemicals.
Many of the receptors important in recognizing fungi and other microbes are multidomain proteins with an extracellular lectin domain. Lectin domains are non-catalytic and are defined by function; they bind ligands, usually sugars, without altering them (De Schutter and Van Damme 2015). Lectins are perhaps most familiar as tools used in cell biology to identify polysaccharidesthroughtheirbinding speci ficities, as when the lectin wheat germ
agglutinin, labeled appropriately and added to fungal cells, reveals through its binding the location of chitin in fungal walls (James and Berbee 2012). In animal and plant cells, receptors bearing different lectin domains can distinguish pathogenic or non-pathogenic microbes by their different cell wall components. Enzymes, in contrast to lectins, bind and alter their substrates. Among enzymes, some, such as chitinases in plants and animals and β-glucanases in insects, have roles both in detecting and in destroying pathogens. 1WhenPlantsandAnimalsFirstMetFungi:InsightsfromtheEvolutionofHostImmuneSystems3
After a host protein recognizes a microbeassociated molecular pattern, host responses usually involve cellular signaling, leading variously to phagocytosis or agglutination of the microbe, to the secretion of antimicrobial toxins, or to programmed cell death that limits a pathogen’s growth by killing its host cell. This chapter does not explore signaling or how a host ultimately disarms a microbe. For reviews that use elegant diagrams to explain host processes, see Erwig and Gow (2016) (mammals); Hatinguais et al. (2020) (mammals); Hoffmann (2003)(Drosophila); Ngou et al. (2022) (plants); and Gauthier et al. (2010) (comparing selected eukaryotes).
Because host receptors and enzymes important in innate immunity are inherited vertically, they can be analyzed phylogenetically. We chose a few of the many host proteins with experimentally demonstrated roles in fungal recognition. Experimental data on the molecular basis for microbial detection come largely from studies of plant models such as Arabidopsis and rice, animal models including mice, and human cell cultures. In the following pages, we discuss the appearance of these proteins through host evolution using published phylogenies where applicable. We then use BLAST searches, comparison of protein domain structures, information on ligand binding sites, and some original phylogenetic analyses to apply newly available data to inferences about when proteins with homologous function may have originated. We use published ages of host clades to infer protein ages, with the caveat that estimates of clade ages vary across studies, especially for unicellular, early-diverging taxa without rich fossil records (Morris et al. 2018; Chang et al. 2015, 2019; Nelsen et al. 2020; Lutzoni et al.
2018; Amses et al. 2022). To specify particular proteins, GenBank accession numbers are given in parentheses in the text or are included in figures.
Immunology as a field has burgeoned, and astounding progress brings with it specialized terminology that can be a barrier to understanding for those from other disciplines. We minimized our use of acronyms and abbreviations, apologizing to colleagues who prefer the more concise notation. Our chapter now focusses on phylogenetic interpretation of proteins with roles in recognizing fungal cell walls.
1.2How Plants Detect and Respond to Fungi
In this section, we speculate on the origins of chitin perception and chitin-degrading enzymes in plants (Fig. 1.1). We review how green algae (chlorophyte and streptophyte algae), bryophytes, and vascular plants perceive fungi, and we explore the diversity in structure and function of plant chitinases the defense-related enzymes that target the fungal cell wall. We present a phylogenetic analysis of one family (glycoside hydrolase 19) of plant chitinases, reconstructing the evolutionary history of this family spanning bacterial, algal, and plant lineages.
1.2.1 Plants Detect Fungi via PatternRecognition Receptors that Contain Extracellular Lysin Motifs (LysM): Lectin Domains that Bind Chitin
The plant immune system allows plants to perceive, initiate defense responses, and mount a resistance to fungal pathogens. Plants evolved to recognize fungi by detecting chitin fragments as microbe-associated molecular patterns. Chitin is an insoluble long-chain polymer of N-acetylglucosamines that can be degraded by chitinases to produce soluble fragments of eight or fewer N-acetylglucosamine residues (Buendia et al. 2018). Other microbe-associated molecular
Fig. 1.1 Overview of distributions of chitin, chitin recognition receptors, and defense-related proteins across lineages that are reviewed over the next pages. Icons created with BioRender.com
patterns perceived by plants include peptidoglycan from bacterial cell walls, a polymer made up of alternating residues of N-acetylglucosamine and N-acetylmuramic acid. In hosts, primary defense responses such as cell wall alterations, production of reactive oxygen species, and expression of defense-related genes like chitinases are initiated by recognition of chitin fragments (Khokhani et al. 2021). A second layer of defense is triggered upon recognition of fungal effectors, which are secreted by some fungi to interfere with plant immune responses, ultimately resulting in hypersensitive responses and programmed cell death, as reviewed by de Wit (2007). The focus of this section will be on primary immune responses to fungi. 4E.D.TrudeauandM.L.Berbee
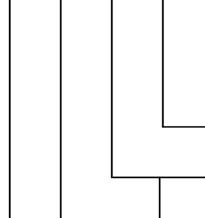
LysM-containing receptors can be receptorlike kinases, with a cytoplasmic kinase domain that can signal when the LysM domains have bound to chitin oligomers (Fig. 1.2a), or receptor-like proteins that lack the cytoplasmic
kinase domain needed for signal transduction (Shiu and Bleecker 2003). LysM domains can detect and bind to not only chitin fragments but also bacterial peptidoglycan. They were first discovered in peptidoglycan-hydrolyzing enzymes, and the name originates from the bacterial cell lysis that results (Buendia et al. 2018; Willmann et al. 2011).
LysM domains of around 40–50 amino acid residues that belong to the carbohydrate-binding module family 50 are found in most organisms, according to the Carbohydrate-Active Enzymes database (www.cazy.org) (Drula et al. 2022). They are not highly conserved in terms of primary sequence, although cysteine pairs separated by one amino acid are found between the three LysM domains of most plant LysM receptor-like kinases and receptor-like proteins, and are required for disulfide bridging between domains (Lefebvre et al. 2012). Secondary and tertiary structures of LysMs are, however, highly
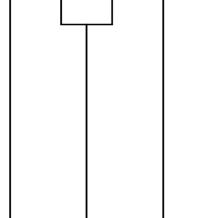
Fig. 1.2 Structure of LysM receptor-like kinases. (a) Domain architecture of CERK1 LysM receptor-like kinase (SP signal peptide, LysM1-3 LysM domains, TM transmembrane domain). (b) Partial alignment of kinase domains in LysM receptor-like kinases from plants and streptophyte algae. Arginine (R) and aspartic acid
conserved, consisting of two α-helices stacked onto two antiparallel β-sheets (Buendia et al. 2018).
1.2.1.1 Chitin Perception in
Land Plants
Typically Involves Complexes of Two or More LysM-Containing Receptors at the Plasma Membrane
These complexes are typically composed of two receptor-like kinases, or one receptor-like kinase and one receptor-like protein. One receptor usually has higher chitin-binding affinity, while the other has an active kinase domain that is necessary for signal transduction (Fig. 1.3). In general, plant LysM-containing receptors have three extracellular LysM domains, with one or more domains binding chitin directly. In Arabidopsis thaliana, chitin oligomers are detected by a receptor complex made up of AtCERK1 and AtLYK5 (Fig. 1.3a). AtLYK5 is a LysM receptor-like kinase, but has an inactive kinase domain, and
(D) residues indicative of RD kinase activity are denoted by an asterisk. Threonine (T) residues essential for chitin signaling are denoted by a number sign. LYK5 from Arabidopsis and LYK3 from Chara lack these residues. Darker shading indicates more highly conserved positions
serves as the high-affinity chitin-binding receptor. AtCERK1 can bind chitin but shows only modest chitin binding compared to AtLYK5. AtCERK1 provides kinase activity and is required for chitintriggered immune responses. Chitin oligomers induce heterodimerization between AtLYK5 and AtCERK1, and AtLYK5 is required for subsequent AtCERK1homodimerization (Fig. 1.3a) (Gong et al. 2020). AtCERK1 phosphorylates the cytoplasmic kinase PBL27, which induces MAPK cascades and immune responses.
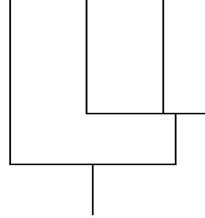
The first plant chitin receptor to be identified— OsCEBiP was cloned from rice in 2006 (Kaku et al. 2006). OsCEBiP is a LysM receptor-like protein that is anchored to the plasma membrane by a glycophosphatidylinositol. It binds chitin but lacks the cytoplasmic kinase domain needed for signal transduction (Jiang et al. 2019). Two CEBiP monomers homodimerize to bind the same chitin oligomer, showing preference for longer chitin fragments consisting of about seven
Fig. 1.3 LysM receptor-like kinases and receptor-like proteins detect chitin and initiate immune responses. (a)
In A. thaliana receptor-like kinases CERK1 and LYK5 form a complex to bind chitin, phosphorylate a cytoplasmic kinase (PBL27), activate a mitogen-activated protein kinase (MAPK) cascade, and induce defense-related gene expression. (b) In rice, O. sativa, CERK1 and receptor-like protein CEBiP form a complex to bind chitin,
phosphorylate a cytoplasmic kinase (RLCK185), activate a MAPK cascade, and induce defense responses. (c) I the moss P. patens, CERK1 activates MAPK signaling cascades. (d, e) Streptophyte algal genomes have CERK1 or LYK5 homologs with untested roles in chitin perception and defense. (d) Chara braunii’s genome encodes seven LysM receptor-like kinases. (e) Nitella mirabilis’s transcriptome encodes a CERK1 homolog
n to eight N-acetylglucosamine residues (Fig. 1.3b).
OsCEBiP then heterodimerizes to form a complex with the LysM receptor-like kinase OsCERK1, which cannot bind chitin but provides
kinase function to the receptor complex (Shimizu et al. 2010). OsCERK1 phosphorylates a receptor-like cytoplasmic kinase (OsRLCK185), which in turn activates the MAPK cascade
resulting in immune responses and defense signaling (Sanchez-Vallet et al. 2014).
In the moss Physcomitrium patens (Fig. 1.3c), PpCERK1, an ortholog to AtCERK1, binds chitin and then initiates MAPK cascade activation (Bressendorff et al. 2016). Physcomitrium patens’s three LYK5 genes are homologs to AtLYK5 (Fig. 1.3a), and silencing of LYK5 genes in P. patens renders plants chitin insensitive (Orr et al. 2020). Additionally, one PpLYK5 was shown to be highly expressed upon infection with the fungus Botrytis cinerea (Reboledo et al. 2021; Orr et al. 2020).
Representing the streptophyte algae, the closest relatives to land plants, the genome of Chara braunii encodes seven LysM receptor-like kinases (LYK1–7) (Fig. 1.3d) and the transcriptome of Nitella mirabilis encodes a homolog to CERK1 from land plants (Fig. 1.3e).
1.2.1.2 LysM Receptor-Like Kinases
Evolved in the Common Ancestor of Land Plants and Streptophyte Algae
Members of the CERK1 clade of LysM receptorlike kinases are as yet known only from streptophyte algae and land plants (Figs. 1.2 and 1.3). Some chlorophyte, red, and glaucophyte algae have LysM domain-containing proteins of uncharacterized function, but proteins that have both the kinase and the LysM domain are as yet unknown in these lineages (Van Holle and Van Damme 2019; Delaux and Schornack 2021). Distantly related LysM domains are also found across bacteria, fungi, and animals, implying that the LysM domain is even more ancient than the divergence of eukaryotes from bacteria, estimated to have occurred over three billion years ago (Battistuzzi et al. 2004; Knoll 2014).
Receptors like CERK1 with active kinase domains are conserved in land plants, with a possible ortholog in the streptophyte alga Nitella mirabilis (Fig. 1.3). Nitella, in Charophyceae, diverged from the ancestor of land plants 635 million years ago (Ma) (estimated from Fig. 1.3 in Morris et al. 2018), suggesting that CERK1 may have been present before plant terrestrialization. That it is highly conserved implies importance for
mediating interactions between land plants and fungi.
For Arabidopsis AtCERK1, mutational analysis showed that kinase function, chitin signaling, and defense-related gene expression required phosphorylation sites at threonine (T) residues T479 and T573 (Fig. 1.2b) (Suzuki et al. 2016). CERK1 bears an RD kinase, having a conserved arginine (R) residue immediately preceding a conserved aspartic acid (D) residue in the catalytic loop of the kinase domain (Greeff et al. 2012). These T, R, and D residues are conserved in CERK1 receptors with active kinases in Arabidopsis (NP_566689), Oryza (XP_015650771), Selaginella (XP_002977121), and Physcomitrium (XP_024388364) (Fig. 1.2b). In the CERK1 homolog from N. mirabilis (JV789971), T, R, and D residues are similarly all conserved.
Arabidopsis LysM receptor-like kinases without kinase activity such as AtLYK5 (AAB80675) lack the homologous T, R, and D residues. Raising questions about functions of its LysM receptor-like kinases, all seven of the genes in C. braunii have a valine (V), not a T that would align at site T573 of AtCERK1, as shown by CbLYK3 (GBG90350) in Fig. 1.2b. All but one of the LysM receptor-like kinase genes in C. braunii lack the R and/or D residues characteristic of RD kinases. Gene family expansion of LysM receptor-like kinases occurred independently in C. braunii and land plants, and future experimental testing of the C. braunii receptors may give fascinating results (Nishiyama et al. 2018). As Nishiyama et al. (2018) point out, the C. braunii receptors’ expansion may have allowed adaption to a range of interactions with chitin-and peptidoglycan-containing organisms.
1.2.1.3 Plant Chitin Receptors Can Initiate Both Immune and Symbiotic Responses
The common ancestor to all land plants may have been preadapted for symbiosis with mycorrhizal fungi, based on the many genes important in fungal colonization that are shared between land plants and the streptophyte algae (Delaux et al.
2015). Since microbial symbionts namely mycorrhizal fungi produce chitin oligomers, perception of these microbe-associated molecular patterns does not inevitably trigger host defense responses. Longer chitin oligomers of six to eight residues can initiate both immune and symbiotic responses but are stronger activators of host immune responses than are short-chain oligomers (Khokhani et al. 2021). The legume Medicago truncatula uses the receptor complex MtLYK9/ MtCERK1 in both its symbiotic and immune responses, and either response can be activated by four-or eight-residue chitin oligomers (Feng et al. 2019). Similarly, in rice, the receptor-like kinase OsCERK1 can detect short-chain oligomers in arbuscular mycorrhizal signaling, suggesting a dual role in immunity and symbiosis (Carotenuto et al. 2017). Discriminating between symbiotic hyphae and pathogenic appressoria may involve additional factors such as plant nutrient status and perception of mechanical stimuli (Khokhani et al. 2021). Evidently, plants are balancing activation of immune signaling and symbiotic signaling to respond appropriately to chitin oligomers.
1.2.2 Lipo-Chitooligosaccharides (LCOs), Chitin-Based Signaling Molecules, Are Widely Distributed Among Fungi
The fungal LCOs modulate plant immune responses when produced by mycorrhizal fungi and nitrogen- fixing bacteria. Like chitin oligomers, LCOs are perceived by LysM receptor kinases that activate downstream signaling cascades (Wanke et al. 2020). First characterized in nitrogen- fixing bacteria that form root nodules in legumes (Roche et al. 1996), LCOs were subsequently shown to initiate signaling for mutualistic symbioses in hosts of arbuscular mycorrhizae (Maillet et al. 2011) and ectomycorrhizae (Cope et al. 2019). Land plant mutualisms via perception of LCOs arose convergently. Arbuscular mycorrhizae are the oldest of the mutualisms with LCO recognition; they involve the earliest-diverging lineages of
land plants and are represented by 407 Ma fossils. Ectomycorrhizae, restricted to seed plants, are known only from 50 Ma fossils, and the rootassociated nitrogen-fixing bacteria, restricted to flowering plants, have a fossil record dating back to ~85 Ma (Bonfante and Requena 2011; van Velzen et al. 2019).
Arbuscular mycorrhizal fungi produce a combination of chitin oligomers and LCOs during colonization of their host plants. Symbiotic outcomes are favored when LCOs suppress the plant immune responses triggered by long-chain chitin oligomers (Maillet et al. 2011). Upon LCO detection, the host initiates signaling pathways that increase the number of lateral roots the most common sites of arbuscular fungal colonization. Many of the signaling proteins are shared by bryophytes and vascular plants (Delaux and Schornack 2021), implying that they were already in place when these clades of land plants diverged ~515–474 Ma (Morris et al. 2018).
Production of LCOs may represent a fungal pre-adaptation to interactions with plants because it is not restricted to mycorrhizal fungi. Rush et al. (2020) showed that fungi in five phyla, including saprotrophs, parasites, and mutualists, produce LCOs. They demonstrated experimentally that LCOs can function as growth regulating hormones in ascomycetes and basidiomycetes (Rush et al. 2020). A role in regulating fungal development may well be ancestral because LCO production, based on its distribution across chytrids, Blastocladiomycota, and Dikarya, originated some 800 Ma (Chang et al. 2019), hundreds of millions of years before land plants.
1.2.3 Plant Perception of Other Fungal Molecules Is Not as Well Characterized
β-1,3-glucan and β-1,6-glucan are other major components of the fungal cell wall, together making up around half of its dry weight. Wanke et al. (2020) found that response to β-glucans, polymers of glucose, varies among plant species and that the degree of polymerization plays a major role in recognition and response. In
Arabidopsis, perception of short-chain β-glucans requires AtCERK1. In contrast in rice, perception of long-chain β-glucans requires OsCERK1 (Wanke et al. 2020).
Arabidopsis can also recognize other fungal microbe-associated molecular patterns, for example producing ethylene in response to xylanases secreted by fungi and bacteria to degrade hemicellulose. Arabidopsis recognizes the fungal endopolygalacturonase enzymes that target pectin in plant cell walls. These molecular patterns are detected at the plasma membrane by receptor-like proteins that contain leucine-rich repeats (Anand et al. 2021; Zeilinger et al. 2016).
Leucine-rich repeats also occur in nucleotidebinding, leucine-rich repeat (NLR) receptors in plants. Well-characterized plant NLR receptors are important in detecting fungal molecules but being cytoplasmic rather than at cell surfaces (Maekawa et al. 2011; Martinon and Tschopp 2005); they are beyond the topics of this chapter. We discuss the roles of leucine-rich repeats in animal toll-like receptors in the upcoming section on detection of fungi by animals.
1.2.4 Chitinases in Plant Defenses Against Fungi
Plants defend themselves against fungi and other microbes by expressing pathogenesis-related proteins including chitinases (Collinge et al. 1993; Pusztahelyi 2018). Plant chitinases compromise fungal cell wall integrity by degrading chitin polymers through the hydrolytic cleavage of glycosidic bonds between β-1,4-linked Nacetylglucosamine monomers (Punja and Zhang 1993). Through this process, chitin oligomers are released, stimulating host immune responses (Sanchez-Vallet et al. 2014). Many plant pathogenic fungi make their first contact with the host in the apoplast. The apoplast, the extracellular space beyond the plasma membrane, contains hydrolytic enzymes and toxins that inhibit fungal growth.
The Carbohydrate-Active Enzymes database (www.cazy.org) classifies plant chitinases into two families: glycoside hydrolases 18 and
19 (GH18 and GH19) (Drula et al. 2022). GH18 and GH19 chitinases show no evidence of homology (Hamel et al. 1997) as they exhibit different catalytic mechanisms and differ in their domain architectures and 3D structures (Cheng et al. 2021). While GH18 chitinases are widely distributed across plant, bacterial, animal, and fungal lineages, GH19 chitinases are mainly found in plants and bacteria (Sharma et al. 2020), although GH19 chitinase function has beenexperimentallycharacterizedin Microsporidia (Han et al. 2016), and using BLAST searches, we found putative GH19 chitinases in some fungi (e.g., GenBank accessions AWI66960 and KAG9575357). Plant chitinases were further divided into seven classes, prior to their classification into GH18 and GH19 families (Fig. 1.4). GH18 includes classes III and V, while GH19 includes classes I, II, IV, VI, and VII (Adrangi and Faramarzi 2013).
Classes were defined based on protein catalytic domains, sequence lengths, and the presence or absence of non-catalytic chitin-binding domains (CBDs) or carbohydrate-binding modules that promote the association of the enzyme with the substrate (Fig. 1.4) (Kitaoku et al. 2022). An alternative framework classifies GH19 chitinases into “loopful” (class I and II) and “loopless” (class IV) chitinases based on the presence or absence of up to six loop regions in the catalytic domain (Hoell et al. 2006; Taira et al. 2011). Recognizing that classes failed to fully reflect phylogeny or capture variation within classes, Orlando et al. (2021) applied a clustering system to divide the catalytic domains of GH19 chitinases into two subfamilies consisting of 51 groups.
There will, however, always be exceptions to the generalized rules involved in classi fication of multi-domain proteins that evolve with domain shuffling. For example, chitinases in a subclass of class III in the horsetail Equisetum arvense and fern Pteris ryukyuensis (Inamine et al. 2015; Onaga and Taira 2008) contain LysM domains. This type of LysM–chitinase conjugate is not usually found in plants, but is common in fungal chitinases (Kitaoku et al. 2022). With BLAST searches, we found a putative GH18 class V
Fig. 1.4 Domain architecture characteristic of classes within GH19 and GH18 chitinases. Classes I, II, IV, VI, and VII are GH19 chitinases that differ in length and presence or absence of a chitin binding domain (CBD), hinge region, and C-terminal extension (Passarinho and de Vries 2002; Orlando et al. 2021). CBDs promote the association of the enzyme with the substrate. Chitinases
chitinase in the spikemoss Selaginella moellendorffii (XP002976579) with a predicted CBD domain more commonly found in class I and IV chitinases of family GH19.
1.2.4.1
Biological Functions and Expression Patterns of Plant Chitinases Vary Among Classes and Species
Since plants do not synthesize chitin, their chitinases can play roles in defense against chitin-containing pathogens such as fungi without compromising their own cellulosic cell walls (Taira et al. 2009). Much research implicates plant chitinases in defense responses against fungi (Kasprzewska 2003; van Loon et al. 2006; Passarinho and de Vries 2002). Some chitinases are synthesized constitutively irrespective of stress conditions, while others can be induced by pathogen infection (Xin et al. 2021). The class I chitinase from rye (Secale cereale BAB18519) is an extracellular inhibitor of fungal growth (Yamagami and Funatsu 1993). Improved
with a C-terminal extension are transmitted to the vacuole, while those without are transmitted extracellularly to the apoplast (Renner and Specht 2012). Classes III and V are GH18 chitinases that share a catalytic domain with a conserved DxDxE sequence motif but show little sequence similarity, and generally lack CBDs (Ohnuma et al. 2011)
resistance to the fungal pathogen Botrytis cinerea resulted when Arabidopsis was transformed with a class IV chitinase from mulberry that is upregulated upon B. cinerea infection (Xin et al. 2021). A class VI chitinase in cashew degraded colloidal chitin and inhibited the growth of the fungus Lasiodiplodia theobromae in vitro by altering hyphal morphology (Oliveira et al. 2020). Class III chitinases from Populus trichocarpa acted on both chitin and chitosan, the deacetylated form of chitin (Zhang et al. 2022).
Many plant chitinases can also contribute to regulating developmental processes such as seed germination and response to abiotic stresses such as freezing and low nutrient conditions (González et al. 2015). A class II chitinase from rye (Secale cereale AAG53610) plays a role in extracellular frost resistance (Yeh et al. 2000). Plant chitinases might be involved in developmental regulation by acting on endogenous arabinogalactan proteins that contain N-acetylglucosamines (van Hengel et al. 2001), but this has not yet been confirmed
experimentally (Hromadova et al. 2021). A role in senescence was suggested when ethylene (a plant hormone associated with senescence) increased expression of a class I chitinase in Arabidopsis (Passarinho and de Vries 2002). An ethylene-dependent signaling pathway was required for induction of this chitinase in plants infected with B. cinerea (Thomma et al. 1999). This class I chitinase might also serve in host defense as it inhibited growth of the fungus Trichoderma reesei in vitro.
1.2.5 Evolutionary History of Plant Chitinases
Since GH18 chitinases can be found across plant, animal, bacterial, and fungal lineages, and since plant GH18 chitinases are more similar to bacterial and fungal GH18 chitinases than other plant chitinases, it follows that GH18 chitinases or “protochitinases” could have been present in the common ancestor of bacteria and eukaryotes. GH19 chitinases, on the other hand, are known mostly from plants and bacteria, although fewer bacterial sequences than plant sequences have been experimentally characterized (Ohno et al. 1996; Sharma et al. 2020; Yano et al. 2021). According to an analysis by Orlando et al. (2021), GH19 chitinases can be divided into a mostly bacterial clade and a mostly plant clade at the basal node of the gene tree. The focus of our analysis is on plant GH19 chitinases, and Orlando et al. (2021) should be consulted for insights into the phylogeny of bacterial GH19 chitinases.
1.2.5.1 Phylogenetic Analysis of GH19 Chitinases
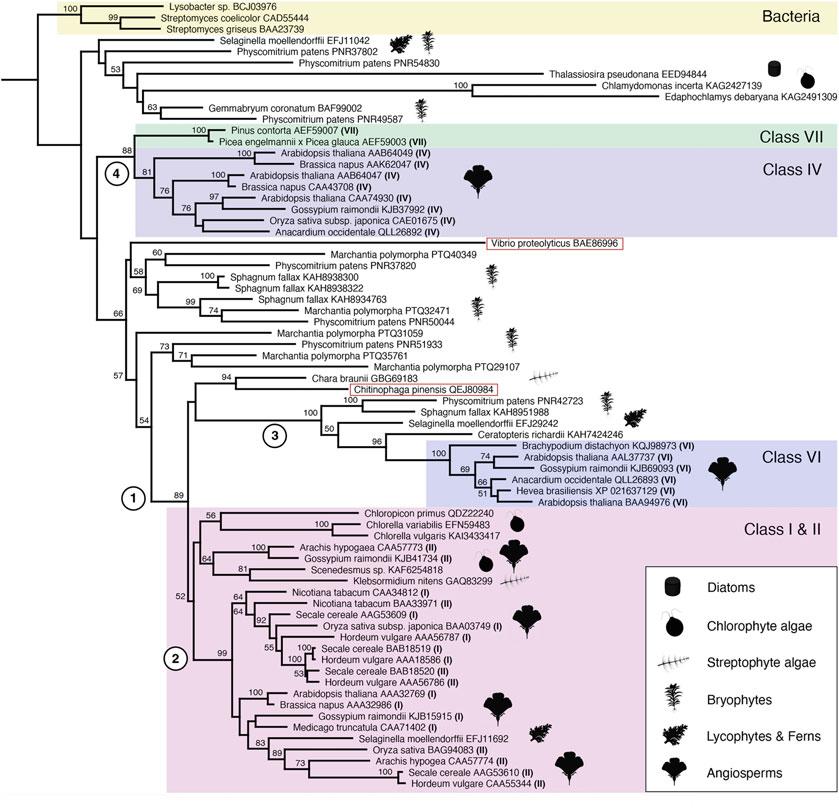
Previous phylogenies of chitinases emphasized seed plants (Taira et al. 2011; Oliveira et al. 2020), but increasingly comprehensive data from chlorophyte algae, streptophyte algae, bryophytes, and seedless vascular plants allow us to explore the deeper origins of GH19 chitinases. To reconstruct the evolutionary history of GH19 chitinases (Fig. 1.5), we analyzed sequences homologous to Arabidopsis chitinases
from land plants, green algae, bacteria, and diatoms. Glaucophyte algae and red algae appeared to lack homologs.
Figure 1.5 clade 1 shows that angiosperm GH19 sequences previously placed in classes I, II, and VI together form a monophyletic group including the seedless vascular plants, bryophytes, streptophyte algae, chlorophyte algae, and a bacterium, Chitinophaga pinensis. Most of these sequences have the chitin-binding sites predicted by Orlando et al. (2021). Some do not, e.g., Secale cereale (AAG53610), and comparison of function of proteins with and without predicted binding sites may be useful in further refining bioinformatic predictions. Denser sampling of bacterial chitinases would be needed to test the signi ficance of their relationship to eukaryotic proteins. Together, class I and class II proteins form the well-supported clade 2 (Fig. 1.5), which also appeared in Oliveira et al.’s(2020) analysis of angiosperm sequences. From our broader sampling, clade 2 sequences appear to have originated before the first divergences of vascular plants 451–431 Ma (Morris et al. 2018), as they encompass flowering plants and Selaginella. Within clade 2, class I and class II sequences are not separated, indicating that chitin-binding domains (Fig. 1.5) are gained or lost convergently through domain shuffling. A good example of lack of monophyly of class I vs class II comes from Secale cereale because one of its class II sequences (BAB18520) is more closely related to its class I sequence (BAB18519), with significant bootstrap support, than it is to another of its class II sequences (AAG53610) (Fig. 1.5).
Our analysis expands the clade of known relatives of class VI chitinases from angiosperms to include other seed plants, bryophytes, and seedless vascular plants, estimated to have diverged starting 515–474 Ma (Morris et al. 2018) (Fig. 1.5, clade 3). Chitinase function has been lost in some angiosperms in class VI, e.g., Arabidopsis (AAL37737) (Fig. 1.5). The chitinases from the seedless plants have retained a glutamic acid residue in the enzyme’s active site that was lost from the corresponding angiosperm proteins, consistent with possible conservation of
Fig. 1.5 GH19 chitinases in plants have origins in the common ancestor of chlorophyte and streptophyte algae, showing gene duplications in early land plants and retention of gene copies. In this maximum likelihood tree from GH19 catalytic domains, species names are followed by GenBank accession numbers, and Roman numerals in parentheses indicate class as assigned by previous literature. Sequences without an associated class number were obtained through BLAST searches of algal, bryophyte,
function of class VI chitinases through early land plant evolution. Classes IV and VII form clade 4 (Fig. 1.5), consisting of seed plants that initially diverged ~365–331 Ma (Morris et al. 2018). We found class IV among flowering plant sequences, and class VII only from gymnosperms, as in Oliveira et al. (Table S2, 2020).
lycophyte, and fern genomes using different classes of Arabidopsis chitinases as queries. The tree was rooted with the bacterial clade as the outgroup. Bacterial sequences nested within plant clades are boxed in red. Bootstrap values greater than 50 are indicated above the nodes. Numbered nodes 1–4 denote well-supported clades that are further discussed in the text. Icons created with BioRender.com
Retention of genes implies continued selection for a diversity of chitinases in land plants, and therefore of defense-related enzymes that target chitin-containing cell walls of organisms like fungi. Our analysis supports the presence of at least one GH19 chitinase in chlorophyte algae and in the common ancestor of streptophyte