Reef health, fish diseases, and habitat connectivity
Puerto Rico Sea Grant Final/Annual Report – October, 2016
A. The Executive Summary
The executive summary covers the entire length of the project and shouldn’t exceed 5000 characters (about two (2) pages). It will be accompanied by the final report narrative. The following must be included:
Title: Reef health, fish diseases, and habitat connectivity: understanding the role of ectoparasites as vectors for disease, energy transfer, and barometers of reef ecosystem health
Summary Prepared: 15 November, 2016
-Name and affiliation of researchers: Paul C. Sikkel, Department of Biological Sciences, Arkansas State University; Amanda W.J. Demopoulos, Southeast Ecological Science Center, US Geological Survey.
Project Number: R-31-2-14
Dates Covered: 15 November 2013 – 15 November 2016
Summary of Impacts and Accomplishments:
1. Objectives:
Objective 1: Further quantify the range and prevalence of fish blood parasites found in the Virgin Islands and Puerto Rico This objective has been completed: A) Blood samples collected from Caribbean damselfish confirm that: 1) blood parasites infect Stegastes but not other damselfish species throughout the US Caribbean (Puerto Rico and USVI), and that, based on molecular evidence, the blood parasite is the same species at all localities. Molecular data also suggest that blood parasites found in Caribbean damselfish and blennies are part of a new Family of apicomplexan blood parasites.; B) Based on morphological analysis, the blood parasite found in yellowtail snapper (Ocyurus chrysurus) is a yeast, the first reported for any marine fish. We are in the process of confirming this with molecular data.
Objective 2: Determine the effects of blood parasites and gnathiid isopods on damselfish and blenny hosts. This objective has been completed as far as possible with the current funding and is being continued with additional funding from other sources (see below). Gnathiid isopods were shown to infect fish as small as settlement size (Fig. 1). Lethal loads of gnathiid isopods for juvenile damselfishes, gobies, and French grunts have been established for multiple size classes (Table 1, Fig. 2). These data were used to establish sublethal loads used to examine sublethal effects on performance (see below). Macrocosms with and without gnathiids have also been established in preparation for additional experiments on sublethal effects of gnathiids. Our data from experiments conducted thus far show that a single gnathiid results in 43-100% mortality for settlement-stage fish, depending on species (Artim et al. 2015, Sellers et al., in
prep.). The number of gnathiids required to induce mortality increases with fish body size. In experiments in which size-matched infected and uninfected Stegastes damselfish were allowed to compete for shelter sites, the uninfected fish won 100% of the time. See below for problems encountered with this objective.
Objective 3: Quantify the contribution of gnathiid isopod parasites to trophic connectivity
This objective has been completed for French grunt, and work is still in progress for brown chromis (see related Objective 4 below). Gnathiid loads on French grunt were quantified from reef and seagrass habitat (Sikkel et al. in press) and from French grunt during the dusk departure and dawn return at multiple sites, using a combination of wild-caught and caged fish. This allowed us to estimate the number of gnathiids and hence the biomass transported during departure and return. We also conducted field experiments demonstrating that gnathiids attached to hosts at departure could remain on the host during transit and therefore be “deposited” in seagrass beds (Fig. 3). On average, for adult-sized fish, an equivalent of 10% of the biomass of each fish is transferred from reef to seagrass via gnathiid isopods, and an average of 7% between seagrass and reef. This constitutes a significant amount trophic transfer. We further demonstrated that French grunt infected by the large parasitic isopod, Anilocra haemuli, typically do not undergo nocturnal migrations (Welicky and Sikkel 2015)
Objective 4: Track the trophic transfer of nutrients from fish hosts to gnathiid isopods to cleaner shrimps using stable isotope enrichment experiments in laboratory and field trials
This objective has been 50% completed and is still in progress. French grunts (N=23) were acclimated to an enclosed saltwater system and provided with a “control” diet at the USGS lab. Once the fish were acclimated to captivity, their food was switched to isotopically labeled freeze dried shrimp enriched with 13C ISOGRO. Initial isotopic analysis of the ISOGRO-labeled diet confirmed that the label was incorporated into the food. The fish were fed the labeled diet for 10 days and then switched back to their control diet for the rest of the study. Fish tissue samples (blood, fin, and scales) were collected immediately prior to switching them to the enriched diet and at the following time points: 0, 8 and 24 hours, 6, 10, 30, and 90 days. Tissue samples were dried, weighed, and analyzed for stable carbon and nitrogen isotopes. We report here the results from the ISOGRO isotopic enrichment study. Results
The preliminary analysis of the ISOGRO enriched diet (freeze dried shrimp) indicated that the label was successfully incorporated into the diet (mean δ13C = +71‰). Isotopic enrichment of the fish tissues first became apparent at t=6h and continued through the 10 days of the enrichment period (Fig. 4, Table 2). At t=30 days, tissues were overall depleted in 13C relative to the t=10 days sampling, but tissues were still enriched relative to the t=0 baseline. By t=90 days the tissues appeared to have returned to their baseline, non-13C enriched, state. Based on these results, we will label the fish for a period of at least 24 hrs prior to the gnathiid labeling experiment.
Advancement of the Field
These data advance our understanding of and capacity to effectively manage Caribbean coral reefs in the following way:
1) By further characterizing the distribution and identity of blood parasites in Caribbean reef fishes, we increase awareness of potential disease-causing micro-organisms in fishes.
2) By demonstrating direct (mortality) and indirect (performance) effects of gnathiid isopods on juvenile coral reef fishes, we demonstrate the potential impact of high levels of gnathiid isopods to negatively impact populations of coral reef fishes.
3) By demonstrating that nocturnal migratory fishes can transport parasitic isopods, we highlight a novel mechanism of energetic transfer between reef-associated habitats, but also expose the possibility that high levels of parasitic isopods on dead reef can be transferred to seagrass beds where they can impact other fish species.
4) Through controlled-diet enrichment studies, we have established a means of ultimately quantifying the transfer of carbon through coral reef ecosystems via external parasites.
2. Problems encountered
Problems were encountered with two of our 4 objectives.
A. Difficulties encountered for Objective 2: We had difficulty finding sufficient variation in natural blood parasite loads in damselfishes and blennies. We therefore focused on examining the direct effects of gnathiids and have initiated a collaboration with Dr. Andrew Campbell at Brown University, an expert on malaria, to amplify blood parasite loads in damselfishes and blennies. No Sea Grant funds were used for components of this collaboration that fall outside of approved Sea Grant activities, but results from Sea Grant supported activities were used to secure additional funding for this objective.
B. Difficulties encountered for Objective 3. There was a long delay in receiving funds allocated to this objective. Thus, the onset was delayed by approximately 1 year. The saltwater system was newly established for this study and, as such, a period of setup and system cycling was required prior to commencement of the study. Initially, wild caught French grunts and Brown chromis were collected from the Florida Keys for use in this study. However, a disease outbreak in the brown chromis resulted in a total loss of that species, so the pilot study was conducted on the French grunts only. We are continuing this project as planned.
3. Research Impacts:
Our work highlights the importance of parasitic organisms in the ecology of US Caribbean coral reefs. Given that gnathiid isopods can directly influence mortality rates and performance of small reef fishes, and given that these organisms are most abundant in dead coral and coral rubble, degradation of coral reefs will likely result in an increase in the abundance of gnathiid isopods and thus impact on fish populations. The finding of multiple varieties of blood parasites has led to further collaborations with investigators working on human blood parasites. This both provides new tools for investigating the impact of blood parasites on Caribbean fishes, but also enables us to use fish blood parasites as model systems to study human diseases.
A. Student participants
1) Joseph Sellers, MS in progress, University of the Virgin Islands: e-mail: joseph.sellers@smail.astate.edu
Role: Lead student, responsible for Objective 2 experiments. Research expenses covered in part by Sea Grant.
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: University of the Virgin Islands: > $24,000.00
Time Period: February 2013 to present
Number of weeks: 202
Number of hours: 8,080
Amounts paid during current reporting period: 0
2) Belytza Velez, BA, University of Puerto Rico.: e-mail: belytza.velez@gmail.com
Role: Undergraduate volunteer: Expenses covered for 6 weeks’ participation.
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: 0 non-federal (student also supported by NSF: $4500.00)
Time Period: April-August 2016.
Number of weeks: 16
Number of hours: 640
Amounts paid during current reporting period: 0
3) Oswaldo Cruz, BA University of Puerto Rico. e-mail: Oswaldo.rullan@upr.edu
Role: Undergraduate volunteer. Expenses covered for 5 weeks’ participation;
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: 0 non-federal. Student also paid by NSF ($3500.00)
Time Period: April – July 2016.
Number of weeks: 12
Number of hours: 480
Amounts paid during current reporting period: 0
4) Nicolas Gomez, BS in Progress, University of Puerto Rico e-mail: Nicolas.gomez@upr.edu
Role: Undergraduate volunteer: Assisted with collection of blood samples from fish.
Cumulative paid by Sea Grant $1000.00
Cumulative paid by Match: CORALations (collaborating partner on Culebra) provided $1500.00 stipend.
Time Period: June-August 2015, April-June 2016.
Number of weeks: 10
Number of hours: 400
Amount paid during current period: $500.00
5) Tina Santos, MS, Silliman University, Philippines. e-mail: tsantos29@gmail.com
Role: Research technician. Expenses covered for 6 weeks in fall of 2015
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: 0 non-federal
Time period: September-November, 2015
Number of Weeks: 12
Number of Hours: 480
Amounts paid during current reporting period: 0
6) Alexandra Hook, BS in progress, Arkansas State University; e-mail: Alexandra.hook@astate.edu
Role: Undergraduate technician and field assistant
Cumulative paid by Sea Grant: $4800.00
Cumulative paid by Match: 0 non-federal. Time period: Summer 2014, 2015, 2016
Number of Weeks: 36
Number of Hours: 1,440
Amounts paid during current reporting period: 0
7) Matthew Nicholson, Ph.D. in progress, Arkansas State University. E-mail: matthew.nicholson@smail.astate.edu
Role: Graduate research assistant. Some field expenses covered during fall of 2015
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: 0 non-federal
Time period: Fall 2015
Number of Weeks: 12
Number of Hours: 440
Amount paid during current reporting period: 0
8) William Jenkins, MS in progress, Arkansas State University. E-mail: wjenkins@gmail.com
Role: Graduate research assistant responsible for Objective 4.
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: 0 non-federal – Salary provided by USGS Time period: Has been conducting the isotope feeding experiments since November, 2015.
Number of weeks: 52
Number of Hours: 2080
Amount paid during current reporting period: 0 Sea Grant
9) Rachel Welicky, Ph.D. completed. Arkansas State University. E-mail: rwelicky@gmail.com
Role: Contributed to Objective 3 experiments. Field expenses covered for 6 months.
Cumulative paid by Sea Grant: 0
Cumulative paid by Match: $15,000.00 from Arkansas State Environmental Sciences Program.
Time period: summer 2014 and 2015
Number of Weeks: 24
Number of Hours: 480
Amount paid during current reporting period: 0 Sea Grant
10) John Artim. Ph.D. completed. Arkansas State University. E-mail: John.artim@smail.astate.edu
Role: Contributed to objective 2 and 3 experiments. Expenses covered for 6 months.
Cumulative paid by Sea Grant
Cumulative paid by Match: $15,000.00 from Arkansas State University Environmental Science Program.
Time Period: summer 2014 and 2015
Number of Weeks: 24
Number of Hours: 480
Amount paid during current reporting period: 0 Sea Grant
B Theses and Dissertations:
The theses linked most closely to the Sea Grant – supported project are those by Joseph Sellers and William Jenkins. Both are still in progress and copies of abstracts will be sent to Sea Grant when they are completed. They and the other students listed above contributed to other aspects of the study and to the publications that include data that were collected during the course of the Sea Grant-supported project. These are listed below.
C. Publications:
Welicky, R.L. Sikkel, P.C. 2015. Decreased movement related to parasite infection in a diel migratory coral reef fish. Behavioral Ecology and Sociobiology. 69: 1437-1446
Artim, J.M., Sellers, J.C., Sikkel, P.C. 2015 Micropredation by gnathiid isopods on settlementstage larval reef fish in the Eastern Caribbean Sea. Bulletin of Marine Science. 91: 479-487
Feature Article
Artim, J.M., Sikkel, P.C., 2016. Comparison of sampling methodologies and estimation of population parameters for a temporary fish ectoparasite. International Journal for Parasitology: Parasites and Wildlife 5: 145-157.
Sikkel, P.C., Welicky R.L., Artim, J.M., McCammon, A.M., Sellers, J.C., Jenkins, W.G., Coile, A.M. (in press). Nocturnal migration reduces exposure to micropredation in a coral reef fish. Bulletin of Marine Science.
Welicky, R.L., Demopoulos, A.W.J., Sikkel, P.C (in press). Stable carbon and nitrogen isotope analyses reveal resource use differences associated with Anilocra spp. parasitism in two coral reef fishes. Marine Ecology
Welicky, R.L., Parkyn, D.L., Sikkel, P.C. (submitted). The energetic effects of large ectoparasitic isopods on two coral reef fish species. Journal of Experimental Marine Biology and Ecology
Renoux, L.P., Dolan, M.C., Smit, N.J., Sikkel, P.C. (accepted pending minor revision). Developing an Apicomplexan DNA barcoding system to detect blood parasites of small coral reef fishes. Journal of Parasitology.
D. Presentations:
1) Blood Parasites of eastern Caribbean Coral Reef Fishes. July 2014, American Society of Parasitologists, New Orleans, LA
2) Ecology of host-parasite interactions on Caribbean coral reefs. April 2015. University of Puerto Rico, Mayaguez and Isla Magueyes Marine Laboratory
3) The potential role of parasitic crustaceans in feeding migrations and trophic connectivity in nocturnal reef fishes. International Congress on Fish Parasites, Valencia Spain, September 2015.
4) Gnathiids at night: Ecology of interactions between mobile ectoparasitic isopods and coral reef fishes. Fish at Night Symposium, Miami, FL, November 2015
5) Low scleractinian coral density is associated with increased density of a common reef fish parasite. International Coral Reef Symposium, Honolulu, HI, June 2016
In addition to these professional presentations, we have given multiple public presentations on St. Thomas, St. John, Puerto Rico, and Arkansas.
E. Other products: A film “Ripple” based on Objective 3 was presented at the Beneath the Waves Film Festival in November of 2015 and won “Best Scientific Merit”.
5. Matching Funds (non Federal)
1) Arkansas State University: $28,000.00 in committed match + Ph.D. student salary ($30,000.00) = $58,000.00
2) Virgin Islands Environmental Research Station: $2,500.00
3) University of the Virgin Islands: $12,000.00 (as student support) + $5,000.00 in user fee reductions.
4) CORALAtions Culebra contributed $2000.00 in student support and supplies.
6. New Extramural funds secured (related to Sea Grant-funded project).
1) Virgin Islands NSF EPSCOR: $14,885.00 (P. Sikkel, PI - awarded) May 2015-May 2016. Project title: Ecology and dynamics of host-parasite interactions in Brewer’s Bay
2) National Science Foundation – Biological Oceanography – $939,000.00 (P. Sikkel, PIAwarded). Project title: Beyond Cleaning Symbiosis: Ecology of “ticks of the sea” on Coral Reefs August 2015 – August 2018.
7. Time and effort of PI and Co-PI:
1) PI Sikkel: Sikkel devoted on average 35% time to the project over the project period. Sea Grant contributed no salary, and Arkansas State University contributed an additional (beyond the normal 9 months salary) $5000.00 in summer salary.
2) Co-PI A. Demopoulos: Demopoulos devoted on average 30% time during the course of the project. Most of her salary was provided by USGS. She received an additional $7,216.00 from Sea Grant funds.
8. Benefits:
1) Our research focuses on a group of organisms (parasites) that dominate biodiversity but that receive very little attention by marine ecologists (outside of aquaculture).
2) Our research has provided valuable information on the role of parasites in Caribbean reef health and trophic connectivity.
3) Our research has provided valuable opportunities for undergraduate and graduate students in the US Caribbean and elsewhere and has contributed directly and indirectly to local US Caribbean economies.
4) Through our community outreach efforts, we have increased awareness among local citizens about the importance and value of coral reefs.
B. Final Report Narrative
1. Research Problem
One of the major challenges facing ecologists, conservationists, and managers of marine reef ecosystems is to understand the complex biological interactions that occur within and between reefassociated habitats and how these interactions are affected by anthropogenic activity. Most research on the diversity and ecology of reef ecosystems focuses on organisms that are easily seen and surveyed by human observers. This occurs in spite of the fact that parasitism is the most common animal lifestyle and thus the most common biological interaction.3,4 Because of their effects on host population dynamics, parasites directly or indirectly influence biodiversity of ecological communities in general. Two aspects of parasite ecology that have received considerable recent attention and seem particularly important to the understanding and management of reef ecosystems include:
1) The role of parasites as vectors of disease-causing organisms, and
2) Their influence on trophic interactions and food-web dynamics. Our previous funding (NSF OCE-121615), enabled us to make progress on the role of parasites by determining which species of fishes in Puerto Rico and the Virgin Islands harbor blood parasites and confirming that gnathiid isopods transmit these parasites. We requested funds from Sea Grant to build on recent results from this first objective and pursue the second, with a focus on external (ecto-) parasites and select host species that undergo daily feeding migrations and thus provide trophic linkages between two or more habitats.
Goals and Objectives: We ultimately aim to develop a comprehensive understanding of the direct and indirect effects of parasites in coral reef ecosystems through their role as disease vectors and influence on trophic interactions and food webs. As a further step toward achieving this ultimate goal, the primary objectives for this proposal are to: 1) Further quantify the range and prevalence of fish blood parasites found in the Virgin Islands and Puerto Rico.; 2) Determine the effects of blood parasites and gnathiid isopods on damselfish and blenny hosts; 3) Quantify the contribution of gnathiid isopod parasites to seagrass-reef and pelagic-reef trophic connectivity via migratory hosts, French grunt, and brown chromis; 4) Track the trophic transfer of nutrients from fish hosts to gnathiid isopods to cleaner shrimps using stable isotope enrichment experiments in laboratory and field trials.
2. Methods
Objective 1. Blood samples were collected from 50-100 Stegastes damselfish from western Puerto Rico and from St. Croix, in addition to supplemental samples from sites where the blood parasites had already been documented. Blood samples were also collected from fish obtained from Curacao. In addition, blood samples from yellowtail snapper (Ocyurus chrysurus) were collected to determine the identity of the blood parasite infecting that species. For each fish, a blood smear was prepared on a glass slide, and additional blood placed in molecular grade ethanol for molecular screening. Each sample was examined via light microscope (after Giemsa staining) and using specifically-designed molecular primers.
Objective 2. Initial efforts focused on expansion of our gnathiid colony and screening of damselfishes and blennies for blood parasites. Expansion efforts were successful, yielding sufficient numbers of gnathiids to conduct proposed experiments. However, blood parasite screening revealed large numbers of fish with low levels of infection, and few with no or heavy infections. We therefore initiated a collaboration with Dr. Andrew Campbell (Brown University) to develop means of enhancing infection levels (these efforts are ongoing and are being funded by grants awarded to A. Campbell and J. Robinson), and focused our experimental efforts on examining direct effects of gnathiids on small reef fishes.
Effects of gnathiids on small reef fishes
Field observations
Lighted plankton traps similar to those used in studies of gnathiids at sites on the Great Barrier Reef (Jones and Grutter 2007) were set between May and August 2014 and 2015 in Greater Lameshur Bay, St. John, US Virgin Islands (18.315024°N, -64.722904°E) a bay dominated by a mix of rocky reef, live and dead Orbicella annularis coral, sand and seagrass. These traps are constructed of PVC and use an LED marker light as a light source and translucent white plastic funnels as one-way entry points. The LED marker light is activated and placed within the trap so that the entry funnel is illuminated. Traps were set two to three hours prior to sunset and were retrieved either three hours after sunset that evening, or three hours post-sunrise the next day. The contents of the traps were filtered through 160 μ plankton mesh and placed in seawater in one or more petri dishes and any gnathiids found were removed, counted and photographed for another study. The traps attracted and collected a wide range of small organisms, including some settlement-stage fishes. For the remainder of the 2014 field season and all of the 2015 season, when we found settlement-stage fish in a sample, the number of attached gnathiids was noted and the fish with attached gnathiids were placed in a separate petri dish for observation or, in one case noted below, the fish and gnathiid were placed in a larger container overnight. In another case, a settlement stage Acanthurus sp. was collected from the water column at night and exposed to three gnathiids in the laboratory overnight.
Fish standard length (SL) and key major-axis dimensions for attached gnathiids were measured from digital microphotographs using ImageJ (Abràmoff et al. 2004). Major-axis dimensions for gnathiids and for their blood and plasma bolus were used to estimate gnathiid juvenile stage and volume of blood and plasma removed by the parasite. Blood and plasma volume was approximated by an ellipsoid whose two minor radii are identical, allowing for estimates from head-on 2-D photographic views. Fish were identified at least to family from the photographic images.
Laboratory experiments
To further assess whether small numbers of gnathiids are capable of causing mortality in settlement-stage fish, we conducted two experiments using newly settled French grunts (Haemulon flavolineatum) collected from Brewers Bay, St. Thomas, US Virgin Islands (18º20'00"N, 64º50'00"W). Fish, ranging from 7-15 mm total length (TL) were collected by escorting them into a plastic container. In the first experiment, 10 fish were randomly assigned to gnathiid-exposed or control treatment. For the former, a total of 48 stage-3 gnathiid zuphea were added to the 1.5 L aerated container of seawater at dusk, and remained with the fish overnight. No gnathiids were added to the control container. The following day, the number of fish that died and number of gnathiids that fed were recorded.
In the second experiment, individual grunts were randomly assigned to 0 (n = 32), 1 (n = 35), 2 (n = 33), or 3 (n = 30) gnathiids placed in 0.40 L containers with a single fish. As before, fish were held overnight and the number of fish that died and number of gnathiids that fed were recorded.
Size-dependent and sublethal effects on hosts
Fish < 30mm FL were held for 5 hours before experimental use while fish >30 mm were held overnight, or 12 hrs before use. This allowed individuals in visibly poor condition to be identified and excluded from experimental use. After the holding period, fish < 30 mm were placed in small cages, and subsequently into plastic urine cups (100 ml capacity) with 50ml of seawater, and a randomly chosen number of juvenile Z3 stage gnathiids (ranging from 0-4 for H. flavolineatum and C. personatus, and 0-10 for S. leucostictus) were introduced to each container. Cages were approximately 5 mm longer and wider than the fish and were used to reduce host predation on juvenile gnathiid (Penfold et al. 2008). Fish >30 mm were similarly caged and added to Tupperware containers with 250ml of seawater and a random number of gnathiids introduced to to each. The baseline number of gnathiids introduced was 0-10 for both damselfish and grunts. However, after it became apparent that S. leucostictus could easily tolerate 10 gnathiids, a range of 0-20 gnathiids was adopted for this species. Fish were exposed to gnathiids overnight for 14-20 h. The number of fish in each trial was dependent on how many could effectively be caught in one day as well as the number of gnathiids obtained from the colony. After each trial was completed, the number of fish that were dead and alive was recorded, as well as the number of gnathiids that fed on each fish. To minimize handling stress, length (FL) was measured after the trial was concluded. To calculate length-weight relationships, additional individuals were collected, measured, and weighed.
The protocol for C. personatus differed slightly from that described above. The small and slender nature of this species allowed for passage through the cage mesh where predation on gnathiids could occur. Because of this, we were only able to collect data for the 10-30mm size range. Also, because of their small size and reluctance to move, it appeared more difficult for gnathiids to find and attach to this species. As with the other species, a random number of gnathiids was selected for each fish. However, in order to ensure micropredation occurred gobies were placed in a container with 10-20 gnathiids and when the number of gnathiids attaching reached the randomly assigned number the goby was removed with gnathiids still attached and placed in their experimental container. This greatly increased the likelihood of gobies being fed on and the possibility of mortality.
Competition Experiments
For the competition trials, juvenile S. leucostictus were collected and contained in the same manner previously described. Measurements such as fork length and weights were collected from each individual. Fish of the same fork length and similar weights were used in competition trials. On average 20 fish would yield approximately 2- 3 pairs that met these standards. Fish were held overnight to ensure they were of even fitness. The number of gnathiids that would cause 35-45% mortality was then applied to one fish randomly and allowed to feed overnight. Both fish were caged during this process. The following morning each fish was removed from the cages. The individual fed on by gnathiids was checked to ensure all gnathiids fed on the fish and that the fish didn’t consume any. A 6L bucket was used as the arena. In the center of the bucket is a circle 7.62 cm in diameter. Within the circle is a 50ml urine cup modified to act as a piece of shelter for the fish. The bottom of the cup was cut off to allow the fish to swim through
it. To facilitate identification, one fish in each pair was randomly selected for marking and the other was sham marked. Approximately 1L of seawater was added to the bucket and the fish were added at the same time. The competition between the fish was recorded for 5 hours. The fish that maintains the territory for the longest period of time is declared the winner of the competition
Objective 3.
To test the assumption that migrating fishes can transport gnathiids between habitats, we conducted an experiment in which 21 French grunt were placed in shallow reef habitat 3 hours before sunset. Seven were retrieved at the time grunts normally depart the reef at dusk, 7 were “swam” for 120 m at the speed migrating grunts typically swim and then removed. A third set of 7 was placed in the seagrass bed and retrieved 3 hours later. Finally, a fourth set of 7 was placed directly in the seagrass bed.
To estimate the average number of gnathiids transported to sea grass habitat during grunt migration, 10-15 grunts were placed in cages near grunt aggregations, and retrieved at the time of departure. This was repeated at 5 sites. Data from previous experiments allow calculation of the number of gnathiids that feed on grunts upon return.
Objective 4.
A pilot study was conducted at USGS-WARC (formally SESC) in Gainesville, FL to estimate the minimum time required for fish tissues (blood, fin and scale) to incorporate an isotopic tracer. The saltwater system was newly established for this study and, as such, a period of setup and system cycling was required prior to commencement of the study. Initially, wild caught French grunts and Brown chromis were collected from the Florida Keys for use in this study. However, a disease outbreak in the brown chromis resulted in a total loss of that species, so the pilot study was conducted on the French grunts only. French grunts (N=23) were acclimated to an enclosed saltwater system and a control diet at the USGS lab. Once the fish were acclimated to captivity, their food was switched to isotopically labeled freeze dried shrimp enriched with 13C ISOGRO. Initial isotopic analysis of the ISOGRO-labeled diet confirmed that the label was incorporated into the food. The fish were fed the labeled diet for 10 days and then switched back to their control diet for the rest of the study. Fish tissue samples (blood, fin, and scales) were collected immediately prior to switching them to the enriched diet and at the following time points: 0, 8 and 24 hours, 6, 10, 30, and 90 days. Tissue samples were dried, weighed, and analyzed for stable carbon and nitrogen isotopes. We report here the results from the ISOGRO isotopic enrichment study.
Results
Objective 1:
Blood parasites were found in Stegastes damselfish collected from all localities, and from both damselfish and blennies from Curacao, suggesting that the blood parasites are widespread in the
eastern Caribbean. Within damselfishes, only members of the genus Stegastes were infected. Molecular data indicate that the blood parasites in Stegastes and blennies are different (Renoux et al. in revision) but that within subfamilies and genera, among locations, the blood parasite is the same. Moreover, molecular data suggest that blood parasites in Stegastes and blennies are members of a new Family of apicomplexan blood parasites (C. Cook, personal comm). The blood parasite from yellowtail snapper appears to be a yeast (C. Cook, personal comm). This is the first yeast ever documented in blood cells of marine fishes, although they have been documented in amphibians.
Objective 2:
Field observations
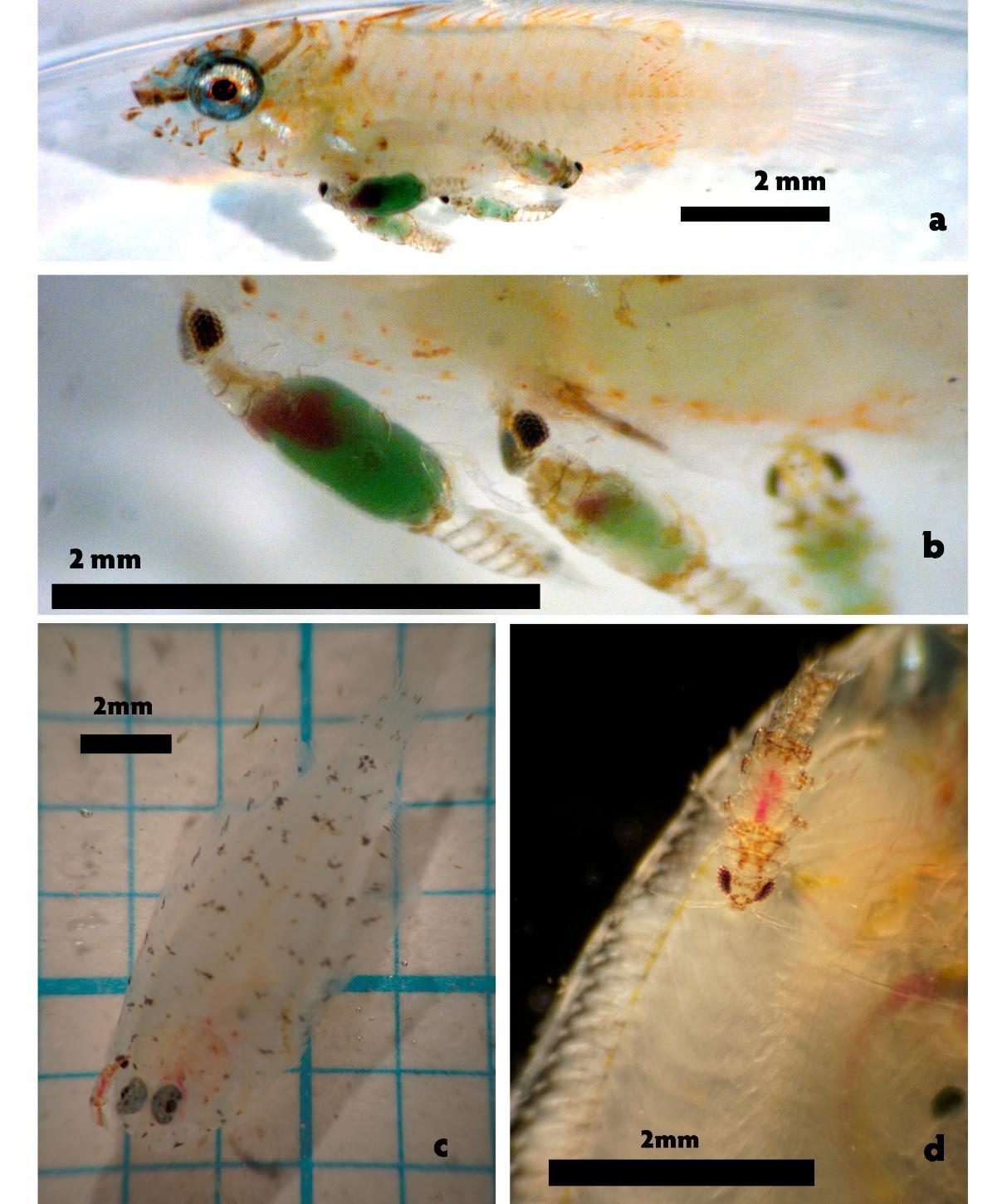
In three instances in 2014, settlement-stage fish removed from traps were infested with gnathiids. The first occasion, in May 2014, was a labrisomid blenny (Starksia sp.; 10.2 mm SL) with five gnathiids (Gnathia marleyi, Farquarson et al. 2012) attached (Fig. 1). This fish and four of the five gnathiids can be seen in figure 1a and 1b. The five gnathiids on this fish varied in size from 1.53 mm to 2.26 mm and extracted an estimated 0.37 μl of blood and plasma volume. Using the blood-volume to body-mass ratio for fish of 0.022 l kg-1 (Schmidt-Nielsen 1997) this 5.01 mg fish had approximately 0.11 μl of blood. Similarly, the whole-body extracellular fluid volume can be estimated using the mean of the measurements Olsen reported for reef-dwelling species 0.189 l kg-1 (Olson 1992). For this fish, the approximation yields a value of 0.95 μl of fluid. The fluids extracted by gnathiids can be seen through the gnathiid carapace in the images in figure 1. These fluids clearly include some whole blood but also appear to include plasma or other extracellular fluid. Our estimate of 0.37 μl of extracted fluids is approximately 39% of this fishes estimated extracellular fluid volume making rapid fluid loss a plausible cause of this larva’s death. One hour of the gnathiid attack on this labrasomid blenny was recorded on video and a short excerpt of the recording, including the point at which the fish’s respiration ceases is included as supplemental content.
The second occasion occurred in June 2014 with the appearance of a larval fish with a single gnathiid attached. This fish was retained in 500 ml of filtered seawater along with the attached gnathiid. The fish was still alive twelve hours later. The image file for this observation was lost no size measurements or fish species identification are available. The third occasion occurred in July 2014 with the appearance of a flounder (Bothus sp.; 14.1 mm SL) with a single gnathiid, approximately 2.13 mm total length. This fish larvae and gnathiid can be seen in the photos in figure 1c and 1d. From size measurements, this appeared to be a secondstage juvenile gnathiid that had extracted approximately 0.06 μl of blood and fluid volume. The total volume of this fish, as measured from photographs, is 12.8 ml. Using the same constants as for the first fish above, we get a blood volume of 0.28 μl and an extracellular fluid volume of 2.42 μl. The 0.06 μl of extracted blood and plasma represent 2.5% of this fish’s estimated extracellular fluid volume a considerably larger proportion than when a similar-sized gnathiid feeds on an adult fish.
In 2015, settlement-stage fish along with gnathiids were found in 52 traps with gnathiids attached to fish in 10 cases. The 10 fish larvae with attached gnathiids included members of the families Gobiidae (n = 6), Apogonidae (n = 1), Bothidae (n = 1), Labrisomidae (n = 1), and Triptyrigiidae (n = 1). In 4 cases, samples were frozen upon retrieval and thus all contents were
dead before the attached gnathiids were discovered. However, in 3 of the remaining 6 cases, the fish was found dead with a single gnathiid attached. The relatively large (25 mm) settlementstage Acanthurus sp was fed on by 2 gnathiids and survived.
Laboratory experiments
In the first experiment, 12 gnathiids fed on the 10 fish exposed to gnathiids (approximately 1 per fish) and all fish died. In contrast, no control fish died. In the second experiment, 20 fed gnathiids were recovered from single-gnathiid trials and in all 20 cases, the fish died. In the 15 cases where the gnathiid did not feed, the fish remained alive. For the two-gnathiid treatment, one or two gnathiids fed on the fish in 15 cases, and in all 15 cases, the fish died. In the remaining 18 cases in which neither gnathiid fed, the fish remained alive. Similarly, for the three-gnathiid treatment, one or more gnathiids fed on the fish in 15 cases and all fish died. In the remaining 15 cases where no gnathiids fed, all fish survived. Finally, in the control treatment where fish were exposed to zero gnathiids, 29 of 30 fish survived.
Size-related mortality comparison for Stegastes
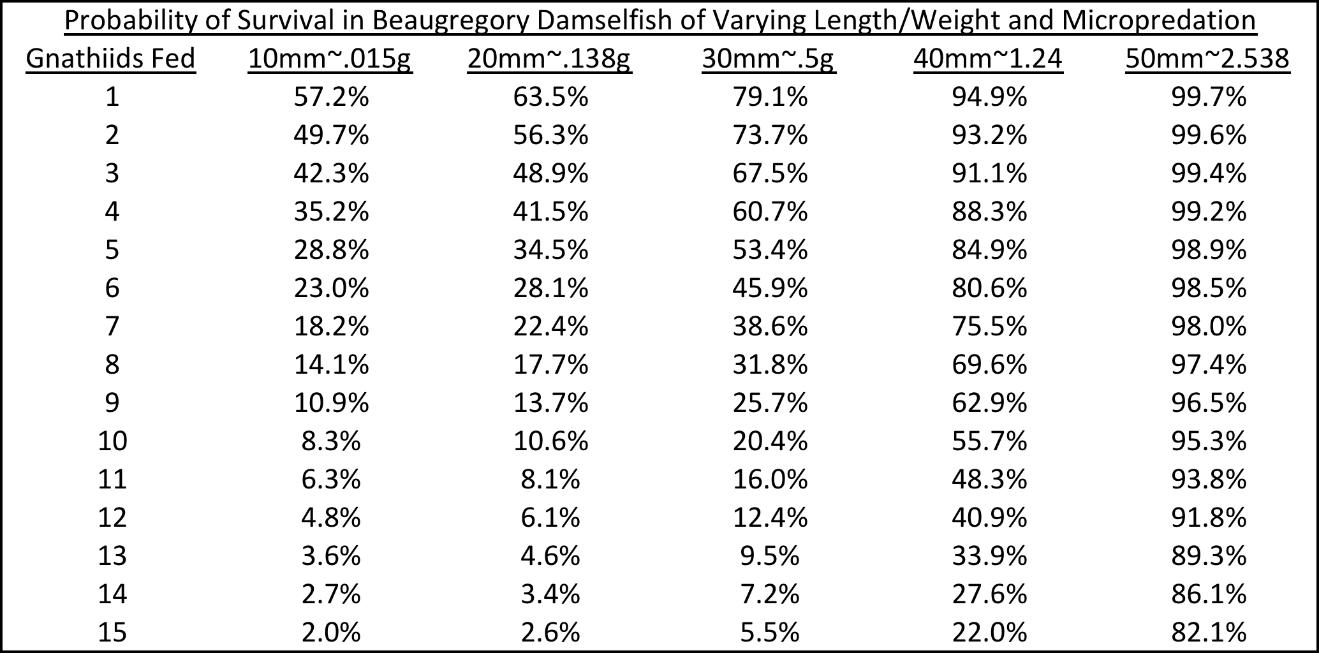
Out of 209 Stegastes leucostictus, 155 were introduced to gnathiids while 54 were controls. Among trials, gnathiids caused mortality in 82 (52.9%) samples while 73 (47.0%) survived micropredation. No mortality occurred in control fish. Predictors of mortality were weight and micropredation. All predictors were significant indicators of survival (P < .001). There were no significant interactions with micropredation and fish size indicating a linear relationship between the variables. The model in Fig. 2 was generated to predict the probability of mortality given a fishes weight and varying micropredation. The probability of survival for various sizes of S. leucostictus is provided in Table 1. The likelihood of survival for a .015-.138g (10-20mm = size-range used in competition experiments) fish when 3 gnathiids feed was 42.3%-48.9%.
Pair-wise competition experiments
Thus far, out of 20 pairwise experiments in which a gnathiid-infected and uninfected fish were allowed to compete, the uninfected fish won in all cases (P < 0.01)
Objective 3
In experiments testing whether grunts can transfer gnathiids between habitats, gnathiid loads varied significantly among treatments (P < 0.01). Loads were highest in fish retrieved immediately at dusk, with about half as many gnathiids on “commuter” fish. Average loads on fish that remained or were placed directly in the seagrass were nearly zero, indicating that grunts can effectively transport gnathiids between habitats (Fig. 3). On average, adult-size grunts transport 7 gnathiids per day from reef to seagrass, or about 10% of their biomass per year.
The preliminary analysis of the ISOGRO enriched diet (freeze dried shrimp) indicated that the label was successfully incorporated into the diet (mean δ13C = +71‰). Isotopic enrichment of the fish tissues first became apparent at t=6h and continued through the 10 days of the enrichment period (Fig. 4, Table 2). At t=30 days, tissues were overall depleted in 13C relative to the t=10 days sampling, but tissues were still enriched relative to the t=0 baseline. By t=90 days the tissues appeared to have returned to their baseline, non-13C enriched, state. Based on these results, we will label the fish for a period of at least 24 hrs prior to the gnathiid labeling experiment.
Discussion of Project Impacts and Findings
Our project is multifaceted and addresses multiple aspects of parasite-host ecology in Caribbean coral reef systems. Our work on blood parasites indicates that they are diverse and widespread and underscore the challenges faced in trying to determine their impact on hosts. Indeed, heavily impacted hosts may die quickly, and fish that are resistant may maintain low levels of infection. More data are clearly needed to understand the dynamics of these infections. As a result of this project, we have been able to generate a collaboration with a malaria biologist and use techniques developed by that lab to better understand blood parasites in Caribbean fishes. We have also trained students and refined molecular techniques to help quantify infections in fishes.
Our work has further demonstrated a direct effect of gnathiids on fish hosts, most surprisingly those of settlement size. Whether through mortality or decreased performance, our findings suggest that gnathiids can have significant impacts on populations of reef fishes. These effects will be linked to the abundance of gnathiids on reefs. The funding we have secured from the National Science Foundation, based on some results from our Sea Grant-funded study is enabling us to address this question. Data are increasingly supporting the hypothesis that live coral is the primary regulator of gnathiid abundance on coral reefs.
While the role of diel migrating fish in trophic transfer has been well established, no previous studies have examined the potential role of parasites. Our data suggest that gnathiid isopods can play a significant role in trophic transfer, fundamentally changing our understanding of how carbon is cycled through coral reef ecosystems. However, because gnathiids can be transported to different habitats, high gnathiid populations on reefs may result in high infestation rates in fish that live in seagrass beds.
Our study is also among the first to go beyond measuring isotope ratios of parasites and hosts and conduct experimental studies to ascertain levels of fractionation between host-parasite and parasite-consumer. We have completed the first phase of our fractionation study (fish) and are continuing with the study. These data will further improve our ability to understand the role of parasites in trophic ecology of Caribbean reefs.
Recommendations
While parasites are an important component of biodiversity, in high numbers they can have significant impacts on host populations. As micropredators, gnathiid isopods in particular can have major impacts on hosts and thus coral reef ecosystems. Given the emerging link between gnathiid populations and live coral, gnathiid isopod populations may be a useful indicator species for reef health. This link also underscores the need to preserve live coral in the US Caribbean.
Artim, J.M., Sellers, J.C., Sikkel, P.C. 2015 Micropredation by gnathiid isopods on settlementstage larval reef fish in the Eastern Caribbean Sea. Bulletin of Marine Science. 91: 479-487
Sikkel, P.C., Welicky R.L., Artim, J.M., McCammon, A.M., Sellers, J.C., Jenkins, W.G., Coile, A.M. (in press). Nocturnal migration reduces exposure to micropredation in a coral reef fish.
Bulletin of Marine Science.
Welicky, R.L. Sikkel, P.C. 2015. Decreased movement related to parasite infection in a diel migratory coral reef fish. Behavioral Ecology and Sociobiology. 69: 1437-1446
Appendices (Tables and Figures)
Table 1. Probability of survival in relation to boby size and level of micropredation in Stegastes.leucostictus.
13C (‰)
Table 2. 13C (‰) in relation to tissue type and time since initial feeding for captive French grunt.
Figure 1. Microphotographs of the larval fish and gnathiids attached to them. (a) Starksia sp. fish larvae with four of the five attached feeding gnathiid larvae with the fifth gnathiid attached on the opposite side of the fish . (b) Closeup of gnathiids attached to the ventral surface of the Starksia sp. larvae. (c) Bothus sp. fish larvae with the attached gnathiid. (d) Closeup of the gnathiid attachment on this Bothus sp. fish.
Fig. 2 Probability of mortality in Stegastes leucostictus in relation to body mass and gnathiid load.
Reef-dusk Commuter Reef-Seagrass Seagrass only
Figure 3 – Results of experiment demonstrating that migrating French grunt can transport gnathiid isopods between habitats. Treatment
4. 13C (‰) in relation to time since onset of feeding trial.