Aire y agua bajo control: el equilibrio indispensable en larvicultura
Los laboratorios de larvas: pilares de la resiliencia en la industria camaronera ecuatoriana
Ecuador alimenta al mundo con su camarón, considerado una superproteína
Del valor nutricional al sabor inigualable de una proteína versátil
El futuro que ellas eligieron
Se reformula el combate a la delincuencia en el Ecuador
Enfermedad de la Postlarva Translúcida: una infección mortal por Vibrio que afecta al camarón de cultivo
Desbloqueo del potencial de la microbiota: el papel del cobre orgánico en la mejora del cultivo saludable del camarón blanco (Penaeus vannamei)
Mioinositol oxigenasa: un actor clave en la lucha del camarón blanco del Pacífico (Penaeus vannamei) contra el estrés por baja salinidad
Efectos de un co-cultivo inactivado por calor de Lactobacillus spp. (postbiótico) sobre el rendimiento de crecimiento y la tolerancia al estrés ambiental en postlarvas de camarón blanco (Penaeus vannamei): síntesis de cuatro ensayos independientes
Técnicas recomendadas para la biorremediación del estuario del Guayas y su área de influencia el perfil costero–marino de la provincia de Santa Elena
Presidente Ejecutivo Ing. José Antonio Camposano
Editora “AquaCultura”
MSc. Shirley Suasnavas ssuasnavas@cna-ecuador.com
Consejo Editorial
MSc. Yahira Piedrahita
PhD. Leonardo Maridueña
Ing. José Antonio Lince Ing. Alex de Wind
Diseño y diagramación Ing. Orly Saltos osaltos@cna-ecuador.com
Ing. Roberto Peñafiel rpenafiel@cna-ecuador.com
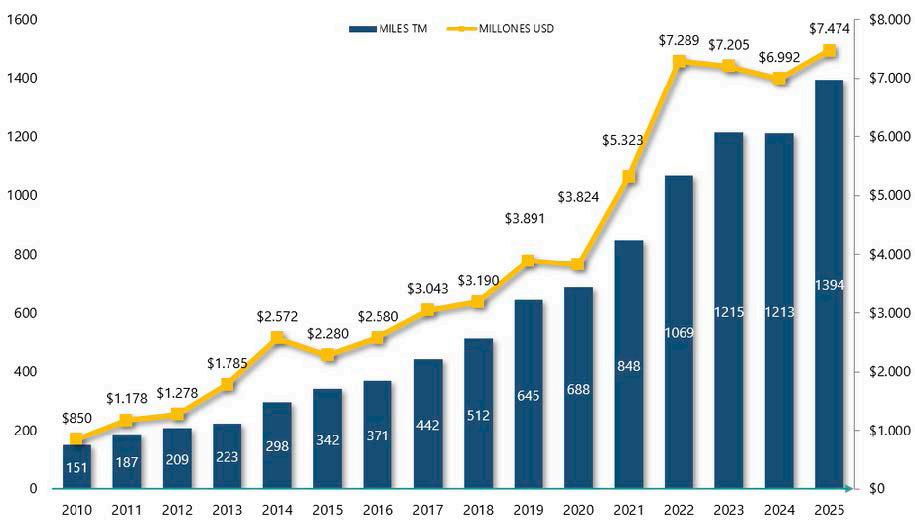
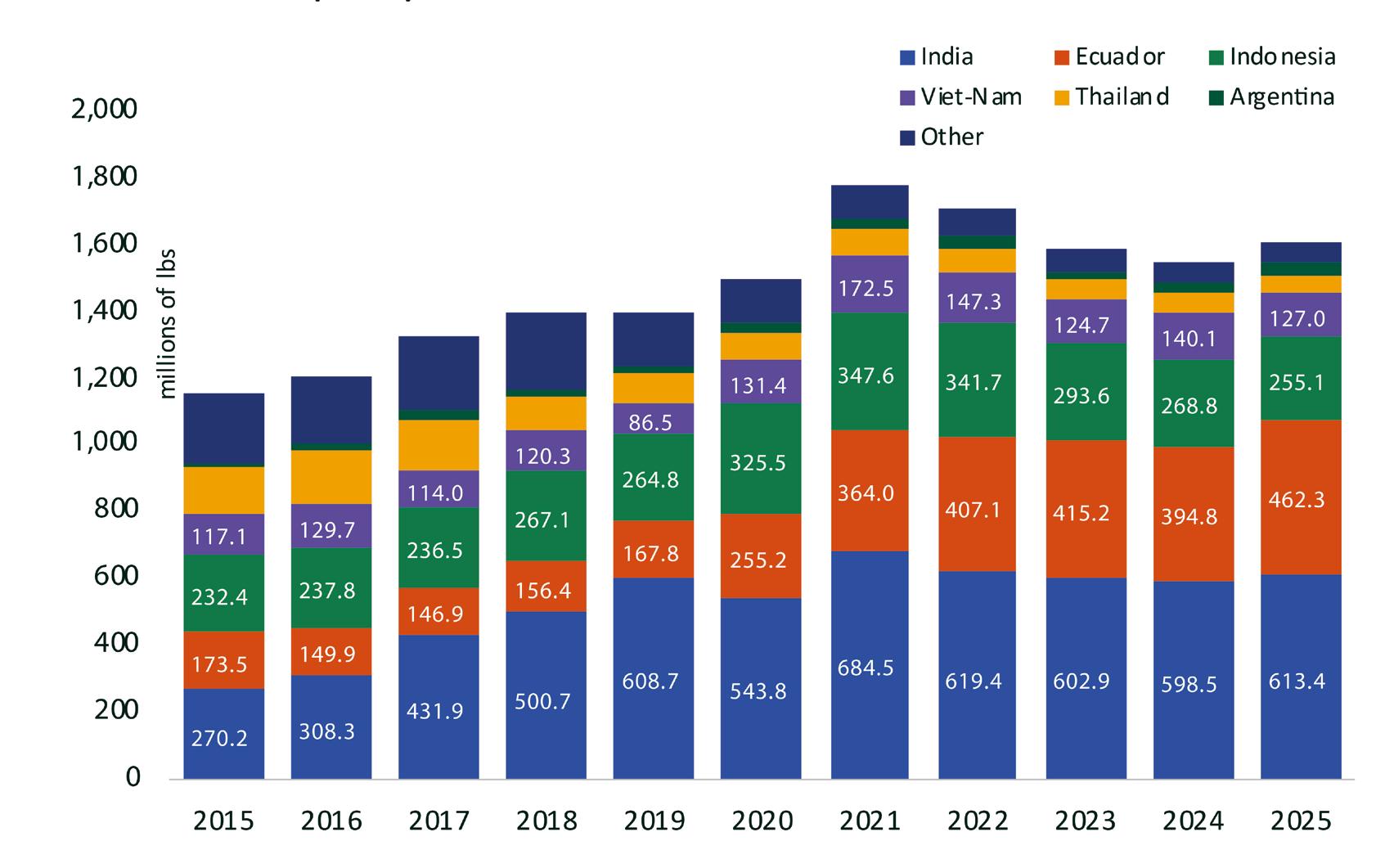
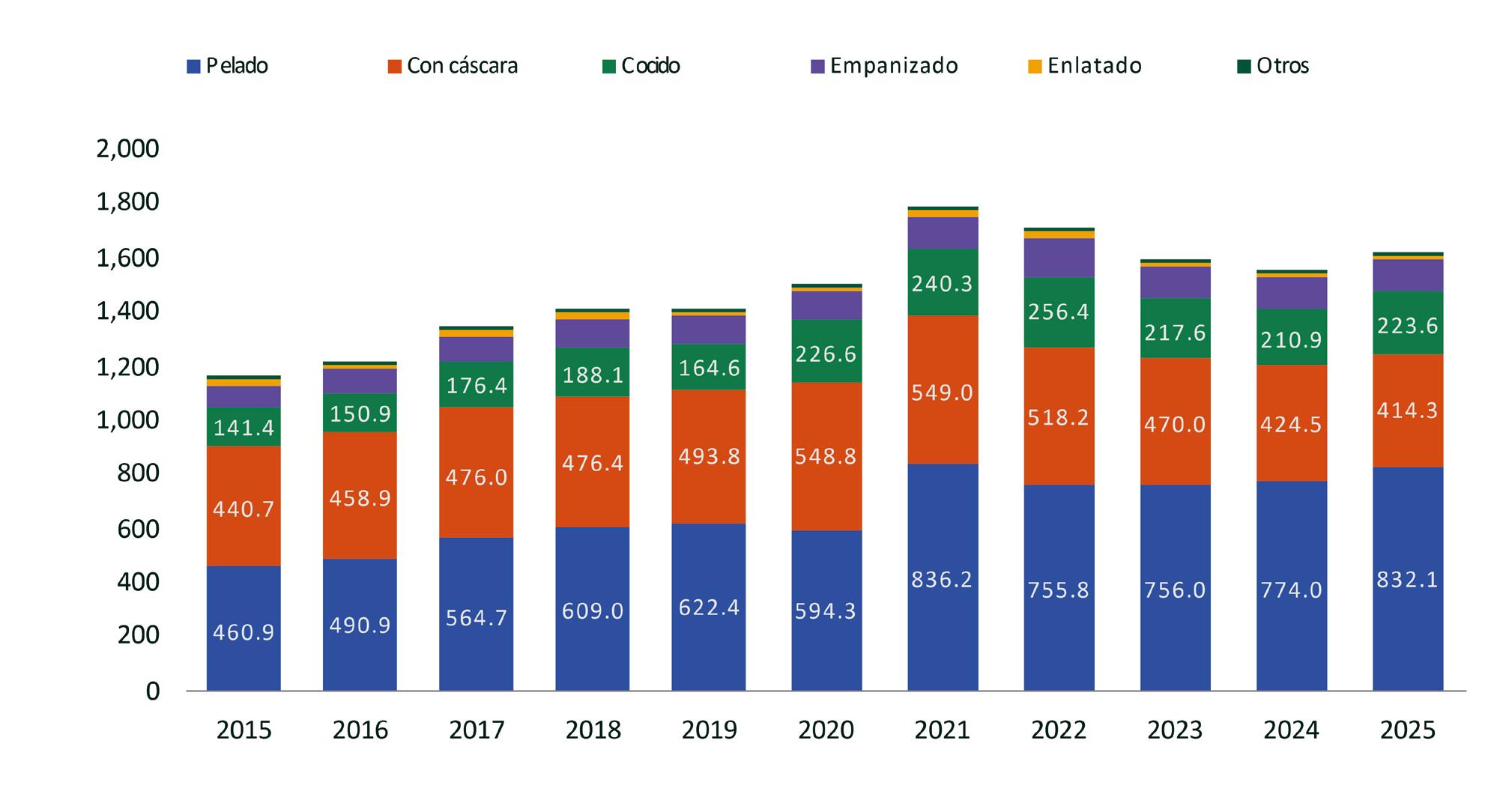
Exportaciones de camarón
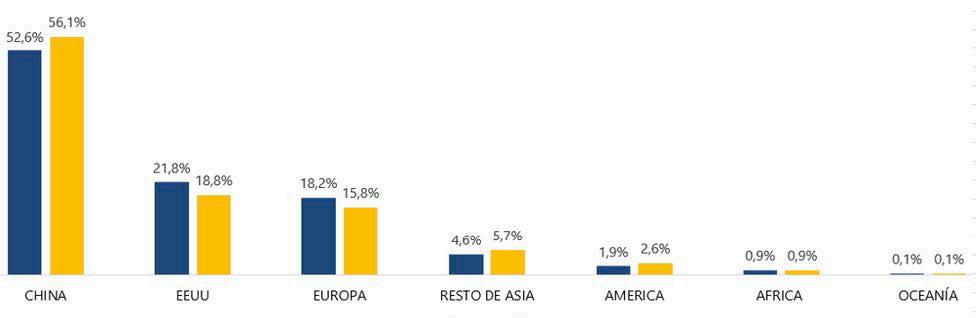
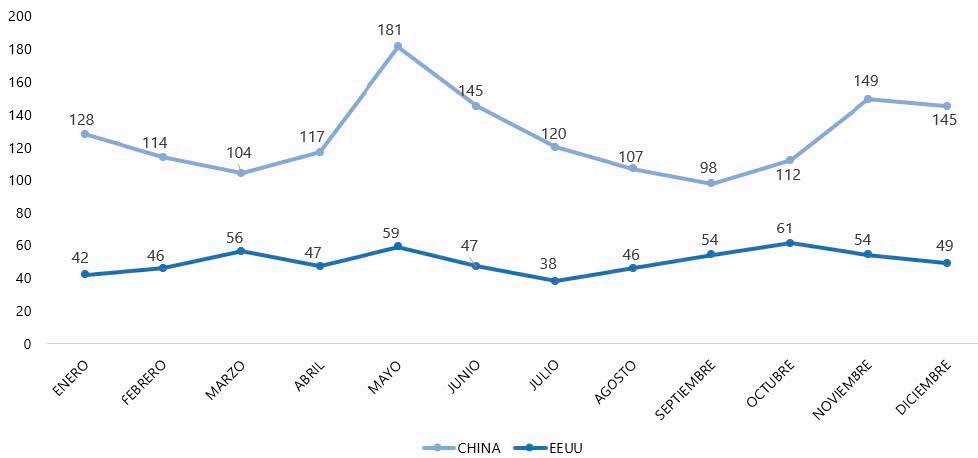
Reporte de mercado de China
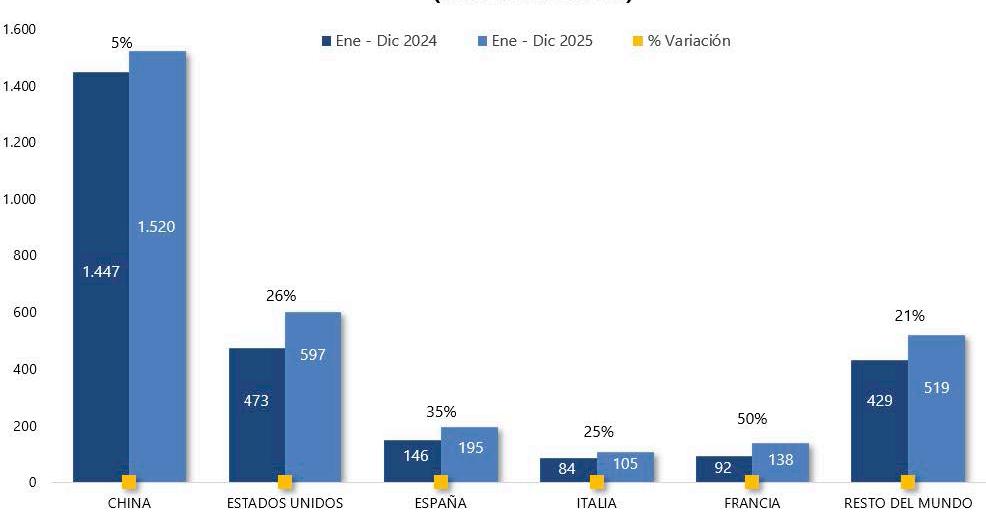
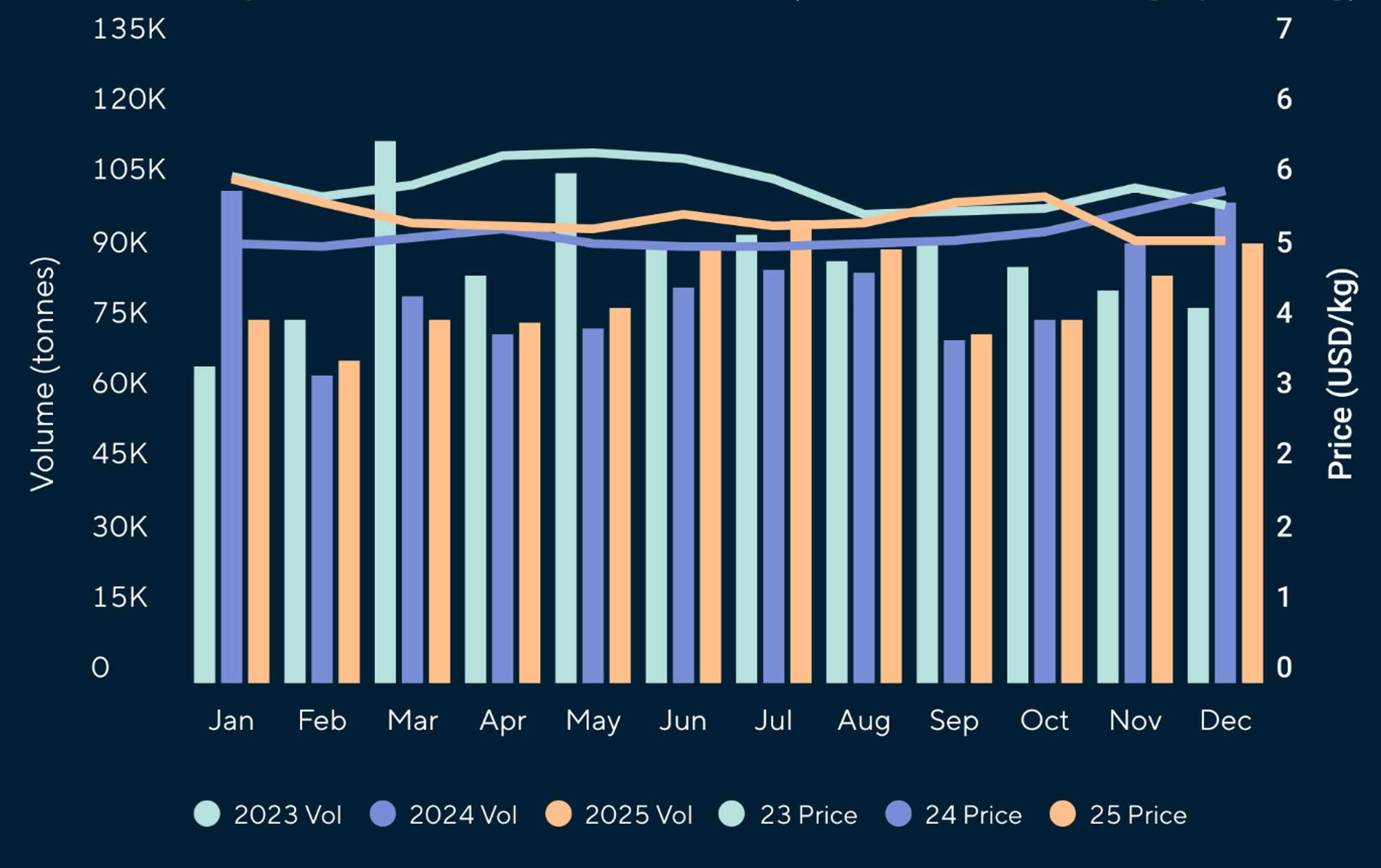
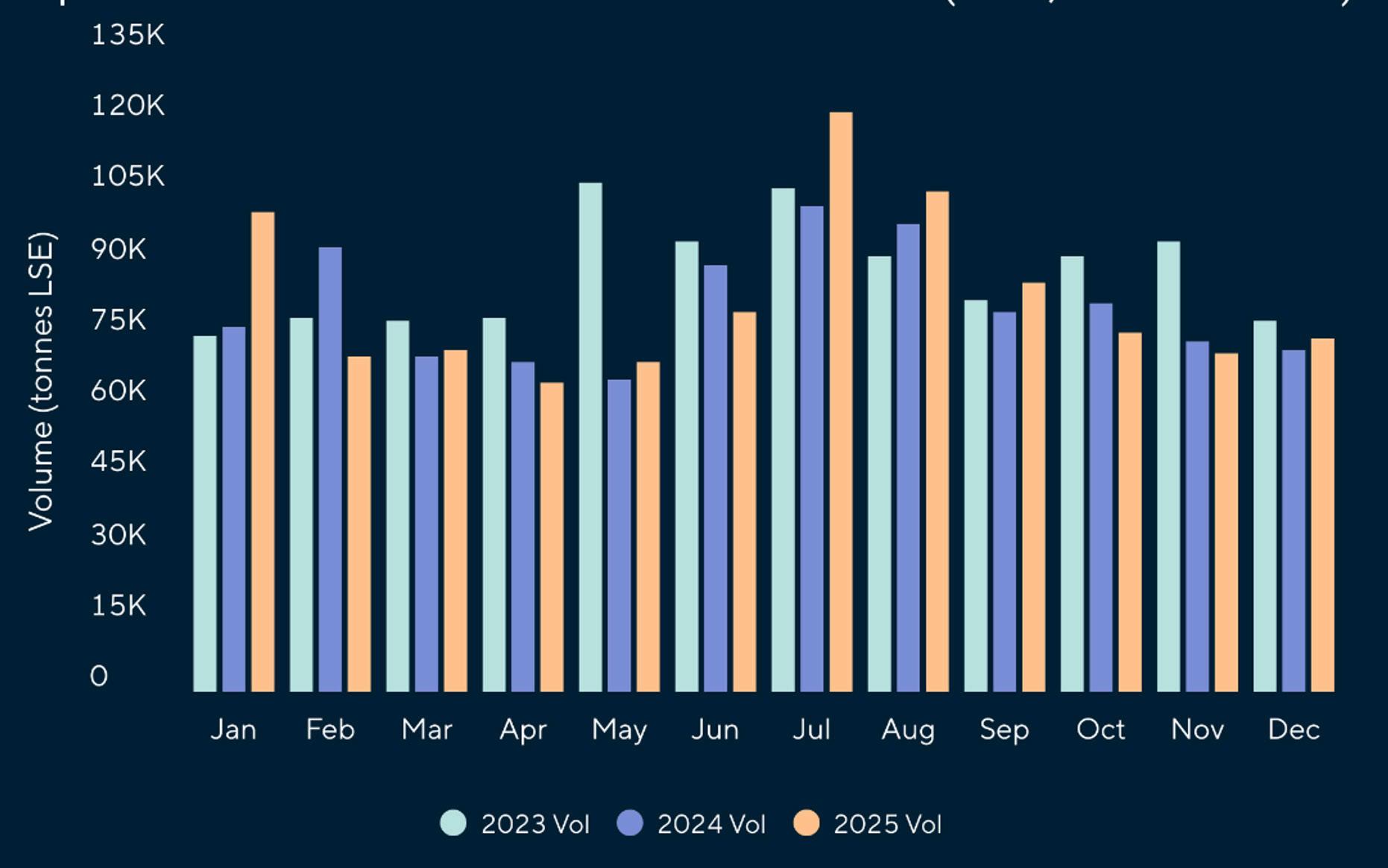
Reporte de mercado de EE. UU.
Noticias del sector
Noticias empresariales
Corrección de estilo
MSc. Daniel Ampuero daniel.ampuero@gmail.com
Comercialización
MSc. Gabriela Nivelo gnivelo@cna-ecuador.com
DJosé Antonio Camposano Presidente Ejecutivo
La sanidad internacional no puede distorsionar el comercio
Para que se produjera un evento sanitario a partir de camarón congelado para consumo tendrían que concurrir múltiples condiciones técnicas improbables relacionadas con supervivencia del agente, exposición y susceptibilidad del hospedero. No reconocer esta diferencia conduce a decisiones desproporcionadas.
urante décadas, el sector camaronero ha cumplido con disciplina las normas sanitarias internacionales, convencido de la necesidad de proteger la producción mundial y de sostener un comercio que funcione sobre bases técnicas claras. Ese compromiso ha sido consistente y responsable.
Sin embargo, la experiencia acumulada en los últimos años obliga a revisar si las normas vigentes reflejan adecuadamente la realidad científica actual. Cuando una enfermedad emergente aparece, su propagación internacional ocurre en plazos muy cortos. Ninguna restricción sanitaria ha logrado impedir ese proceso, porque la biología y la dinámica global no se detienen en las fronteras. Lo que sí ha ocurrido es que determinadas exigencias sanitarias han terminado utilizándose para impedir comercio sin que exista una relación proporcional con el riesgo real.
El caso del IHHNV resulta ilustrativo. La evidencia científica disponible demuestra que, en las condiciones productivas actuales, este virus no genera impactos económicos significativos y tampoco se han documentado efectos relevantes en poblaciones silvestres. Además, estudios han confirmado que fragmentos de su material genético ya están integrados en el genoma de muchas poblaciones de camarón en el mundo.
Esta realidad tiene implicaciones prácticas. Los métodos de detección molecular, como el PCR, pueden identificar ese material genético aun cuando no exista infección activa ni capacidad real de transmisión. En otras palabras,
puede detectarse material genético sin que exista enfermedad ni un riesgo sanitario efectivo.
A partir de allí, resulta indispensable diferenciar escenarios. El riesgo epidemiológico asociado a animales vivos destinados a cultivo no es comparable con el de camarón congelado destinado a consumo humano. Un organismo vivo puede replicar y transmitir un agente infeccioso dentro de un sistema productivo; un producto procesado y congelado carece de esa capacidad biológica. Para que se produjera un evento sanitario a partir de camarón congelado para consumo tendrían que concurrir múltiples condiciones técnicas improbables relacionadas con supervivencia del agente, exposición y susceptibilidad del hospedero. No reconocer esta diferencia conduce a decisiones desproporcionadas.
La gestión sanitaria moderna debe sustentarse en análisis de riesgo claros, diferenciados y basados en evidencia actualizada. No se trata de debilitar controles ni de cuestionar la importancia de la sanidad animal, sino de asegurar que las medidas respondan a riesgos reales y no se conviertan, por interpretación o diseño, en obstáculos injustificados al comercio.
El comercio internacional requiere reglas previsibles y coherentes con la ciencia disponible. Cuando la evidencia evoluciona, las normas deben evolucionar con ella. De lo contrario, se debilita no solo la sanidad, sino también la confianza en las reglas que sostienen el comercio internacional•
DIRECTORIO
PRIMER VICEPRESIDENTE
Ing. Luis Francisco Burgos
Ing. Ricardo Solá
Dr. Alejandro Aguayo
Ing. Chris Olsen
Ing. Francisco Pons
Ing. José Antonio Lince
Ing. Jorge Redrovan
Ing. Alex de Wind
Ing. Kléber Siguenza
Ing. Rodrigo Vélez
Ing. Iván Rodríguez
Ing. Juan Carlos Vanoni
Ing. Alejandro Ruiz-Cámara
PRESIDENTE DEL DIRECTORIO
Ing. Marcelo Vélez
VOCALES
Econ. Heinz Grunauer
Ing. Víctor Ramos
Ing. David Eguiguren
Ing. Humberto Dieguez
Ing. Eduardo Seminario
Ing. Miguel Uscocovich
Ing. Vinicio Aray Dueñas
Econ. Sandro Coglitore
Ing. Rodrigo Laniado
Ing. Roberto Aguirre
Blgo. Carlos Sánchez
Ing. Diego Puente
SEGUNDO VICEPRESIDENTE
Ing. Fabricio Vargas
Ing. Johnny Adum
Sra. Verónica Dueñas
Ing. Alex Elghoul
Ing. Bastien Hurtado
Ing. Luis Burgos
Econ. Wolfgang Harten
Jorge Gonzalez
Andres Rivadulla
Ing. Héctor Marriott
Ing. Edison Brito
AIRE Y AGUA BAJO CONTROL: EL EQUILIBRIO INDISPENSABLE EN LARVICULTURA
Durante las primeras etapas del desarrollo larvario, cuando la bioseguridad es más imprescindible, el manejo del aire ambiental, adquiere la misma relevancia que el tratamiento del agua. Su control permite reducir riesgos sanitarios, limitar fuentes de contaminación y sostener condiciones ambientales estables. No es un complemento operativo: es parte del núcleo del proceso productivo.
Esa visión integral es la que define el trabajo del laboratorio de larvas de Promarisco ubicado en Chanduy, provincia de Santa Elena, y cuya experiencia analizaremos en el presente artículo. Según explica Gustavo García, Director de Larvicultura y cultivo Vannamei, este enfoque ha sido clave para que el laboratorio sea reconocido dentro del programa Sustainable Shrimp Partnership (SSP), una iniciativa que distingue a los centros que demuestran control operativo y cumplimiento de estándares aplicados a la producción larvaria. El reconocimiento no se apoya en un solo proceso, sino en la coherencia entre las prácticas productivas, el manejo ambiental y la forma en que se gobiernan los factores que sostienen la vida desde su etapa más temprana.
A partir de este aval, el equipo de la revista AQUACULTURA se propuso conocer de cerca las acciones implementadas en este laboratorio, con el objetivo de comprender cómo el control ambiental, desde el aire hasta el agua, se integra en la gestión diaria y contribuye a sostener procesos productivos consistentes desde el origen.
“En larvicultura, el control no empieza cuando la larva entra al tanque, empieza mucho antes. Aire y agua son variables que no se pueden separar, porque cualquier desequilibrio en una de ellas termina afectando todo el sistema. Nuestro trabajo consiste en anticiparnos, en crear condiciones estables desde el primer momento, porque ahí es donde se define la sanidad de todo lo que viene después”.
Gustavo
García Director de Larvicultura y Cultivo Vannamei
Un laboratorio que nació en medio de una crisis
En 2010, cuando la Mancha Blanca se había convertido en una preocupación constante para la industria camaronera, en Chanduy comenzó a tomar forma una iniciativa pensada para actuar desde el origen del problema. Lejos de los centros habituales de producción, el laboratorio fue concebido como un espacio de control sanitario, enfocado en la multiplicación de camarones libres de patógenos específicos (SPF) y en la provisión de larvas sanas para las fincas.
En esta caso no se trató de ampliar la producción, sino de intervenir en puntos críticos del ciclo. La ubicación aislada y el diseño del proyecto respondían a esa lógica desde el inicio.
Una base genética cuidadosamente construida
El arranque fue preciso. Se seleccionaron 500 animales certificados como libres de enfermedades incluidas en la lista de la OIE. A partir de ese grupo inicial se estableció un esquema de replicación mensual que, con el tiempo, permitió conformar una población cercana a los 4,000 reproductores que eran enviados al laboratorio de maduración en San Pablo.
El control sanitario no quedó solo en manos del laboratorio. Cada año, los animales eran analizados y certificados por la Universidad de Zaragoza, un respaldo externo que aportaba trazabilidad y sostenía los estándares de bioseguridad sobre los que se apoyaba toda la operación.
Cuando un cierre marca un inicio
Este modelo se mantuvo durante casi una década, pero en 2019, un brote del síndrome de Mortalidad Temprana (EMS) en el laboratorio de Mar Bravo marcó el cierre del proyecto original. El hecho obligó a detener la operación, pero no borró lo construido.
Las instalaciones de Chanduy quedaron
inactivas, aunque conservaban atributos que pronto cobrarían un nuevo sentido: aislamiento geográfico, experiencia acumulada en manejo sanitario y una certificación SPF previa. Sobre esa base, el espacio fue reconvertido en un hatchery de larvas con mayor capacidad y eficiencia, dando inicio a una nueva etapa.
La transición no fue solo operativa. Marcó el comienzo de un proceso de transformación que redefiniría la escala, el enfoque y la forma de organizar el laboratorio, sentando las bases del modelo que hoy estructura su funcionamiento.
Actualmente, el laboratorio es reconocido no solo por su producción larvaria, sino por una decisión que marca la diferencia: contar con una planta de tratamiento de aguas integrada a su operación. En un laboratorio de larvas, el agua no es un recurso estático: se transforma a lo largo del proceso productivo, acumulando carga orgánica, bacterias y residuos que, sin un manejo adecuado, pueden convertirse en un riesgo sanitario y ambiental.
La planta de tratamiento permite cerrar ese ciclo y mantener el control en cada etapa, acompañando el recorrido del agua desde que sale de producción hasta su descarga final, y asegurando que los efluentes no regresen al sistema ni impacten el entorno. En términos concretos, tratar el agua es una forma de proteger simultáneamente a
la larva, al laboratorio y al ambiente que lo rodea.
El agua en movimiento: asegurar el suministro
El sistema productivo inicia en una estación de bombeo que garantiza el suministro constante de agua a las salas de producción. Bajo la supervisión de Carlos Mujica, el laboratorio opera tres bombas de 7.5 HP que captan el agua a través de un sistema de filtración inicial.
Es el sistema principal del abastecimiento de agua de mar para abastecer ininterrumpidamente de agua salada al laboratorio, almacenada en dos mega reservorios de 1,000 tns. cada uno, y es distribuida a los diferentes reservorios de cada sala para los distintos usos de recambios y procesos en la producción de larvas.
“El sistema de bombeo es el que garantiza que todo funcione sin interrupciones. Mantener un suministro constante y estable de agua es fundamental para que los recambios se hagan cuando corresponde y bajo las condiciones adecuadas. La continuidad operativa empieza por asegurar que el recurso llegue siempre en las mismas condiciones”.
Carlos Mujica Jefe de Mantenimiento
Tratamiento en funcionamiento
El impacto del sistema se percibe más allá del laboratorio. La eliminación de malos olores y focos de contaminación ha permitido que en el canal de drenaje crecer el mangle y otras plantas nativas, evidenciando una mejora en la calidad del agua descargada y el control en el manejo de residuos sólidos.
Este modelo solo fue posible gracias a la disponibilidad de un espacio amplio. El predio de 50 hectáreas, con un área específica destinada al tratamiento, permitió integrar la planta sin interferir con la operación productiva, algo que no habría sido viable en la ubicación anterior del laboratorio.
Así, la planta de tratamiento deja de ser una infraestructura secundaria y se convierte en el punto donde la bioseguridad se completa. No termina en el tanque ni en la sala de producción: se extiende hasta la descarga final, protegiendo al mismo tiempo a la larva, al laboratorio y al ambiente acuático que lo rodea.
¿Cómo funciona el sistema de tratamiento?
El tratamiento de las aguas residuales se desarrolla en tres fases consecutivas, diseñadas para acompañar el recorrido del efluente de manera progresiva y controlada.
Fase 1: concentración y sedimentación
En la primera etapa, todas las aguas provenientes de la producción y de la limpieza del laboratorio confluyen en un recolector central. Allí se aplican biorremediadores naturales inorgánicos que facilitan la
sedimentación de sólidos y desechos, reduciendo la carga inicial del efluente antes de su paso a la siguiente fase.
Fase 2: filtración y depuración
Una vez sedimentada, el agua avanza hacia la zona conocida como el laberinto. En este tramo, el efluente atraviesa un sistema de filtración compuesto por mallas de distintos micrajes y carbón activado. Este proceso permite retener partículas remanentes y absorber residuos químicos, afinando el tratamiento antes de la etapa final.
Fase 3: sedimentación final y disposición responsable
La última fase se desarrolla en piscinas de sedimentación, donde el agua recibe un nuevo tratamiento con biorremediadores.
A partir de aquí, el flujo se divide: el agua decantada es rebombeada hacia el canal
de drenaje, mientras que los sedimentos se secan y son entregados a un gestor autorizado, completando el manejo responsable del residuo.
Donde se vigila lo invisible: control de calidad y bioseguridad
Más allá de los tanques y las salas de producción, el control del laboratorio se sostiene en áreas donde el trabajo no siempre se ve, pero define cada decisión operativa. Los laboratorios de análisis funcionan como un sistema de alerta temprana, encargado de monitorear lo que ocurre en el agua, en las larvas y en cada insumo que ingresa al proceso.
Bacteriología: leer el comportamiento del sistema
El laboratorio de bacteriología, a cargo
Sistema de bombeo
Fase 1: concentración y sedimentación Fase 2: filtración y depuración
Fase 3: sedimentación final y disposición responsable
“La bacteriología nos permite leer lo que el sistema está diciendo todos los días. No se trata solo de detectar bacterias, sino de entender su comportamiento en el tiempo. Comparar resultados, ver tendencias y actuar a tiempo es clave para evitar que un cambio pequeño se convierta en un problema mayor dentro del laboratorio”.
Kattiana
Ricardo
Jefe de Patología y Sanidad Animal
de Katiana Ricardo, realiza un monitoreo diario y continuo de todas las muestras críticas: agua, nauplios, larvas, algas y artemia. Las muestras son analizadas en diferentes tipos de agares como agar marino, TCBS, Chromagar vibrios, Chromagar pseudomonas y Saboroud, una técnica que permite identificar la presencia de colonias bacterianas potencialmente patógenas.
Los resultados se obtienen en un plazo de 24 horas y se comparan con una base de datos histórica del laboratorio. Esta lectura en contexto permite detectar incrementos bacteriológicos —especialmente frecuentes en temporada invernal— y anticiparse a posibles desequilibrios. Cuando es necesario, se activan intervenciones puntuales mediante tratamientos y probióticos, ajustados a los niveles detectados.
PCR: confirmar lo que no se ve
Mientras la bacteriología observa el comportamiento microbiano, el laboratorio de PCR, liderado por Henry Bayona, se enfoca en la detección específica de patógenos de ADN y ARN que afectan al camarón. Entre ellos se incluyen agentes asociados a enfermedades como WSSV, IHHN, NHP.
El proceso abarca desde la extracción del material genético y la medición de concentraciones, hasta la amplificación en termocicladores con capacidad para hasta 96 muestras simultáneas. El análisis final se realiza mediante electroforesis en gel de agarosa, permitiendo confirmar la presencia o ausencia de patógenos.
“El PCR nos da la confirmación que no se puede ver a simple vista. Cada muestra que analizamos es una herramienta para tomar decisiones con respaldo, no con suposiciones. Detectar a tiempo un patógeno protege no solo a la larva, sino a toda la cadena productiva que depende de ella”.
Henry Bayona Subjefe de Biología Molecular
Los resultados están disponibles entre 24 y 48 horas e incluyen el monitoreo de larvas despachadas, nauplios entrantes y muestras externas de terceros. La técnica aplicada cuenta con aval de la Universidad de Arizona, validado a través de la prueba RING TEST anual. Además, el laboratorio analiza camarones de finca y muestras de plantas procesadoras para detectar genes patógenos de interés humano, como vibrios y citotoxinas, requeridos para procesos de exportación. Toda la infraestructura está diseñada para evitar la contaminación cruzada entre muestras.
Sostenibilidad: ordenar los residuos para reducir el impacto El control no termina en el análisis sanitario.
La gestión de residuos forma parte del mismo enfoque integral. Bajo la coordinación de Kenya Guerra, el laboratorio implementa un sistema de segregación de plásticos reciclables y no reciclables, que son entregados a recicladores autorizados.
Los desechos peligrosos, como aceites y filtros, son gestionados por entidades autorizadas por el Ministerio del Ambiente, asegurando su disposición final, reutilización o reciclaje, según corresponda. A la par, se han incorporado medidas para reducir el uso de plásticos de un solo uso, como la compra de insumos en envases de mayor volumen y el uso de fundas reciclables para los medios de cultivo. Estas prácticas han posicionado al laboratorio entre los centros de producción larvaria con mayores estándares de sostenibilidad en el país.
Kattiana Ricardo, responsable del área de bacteriología
“La sostenibilidad no es un área separada del proceso productivo: es parte de la misma lógica de control. Gestionar los residuos, reducir el uso de plásticos y manejar adecuadamente los desechos peligrosos es una forma de cerrar el ciclo y de asumir la responsabilidad sobre lo que generamos”.
Kenya Guerra
Jefe
de Sostenibilidad y Control de procesos
Personas, liderazgo y cultura operativa
Detrás de cada área técnica hay un componente humano que sostiene el funcionamiento del laboratorio. La selección de personal capacitado, la motivación constante y el liderazgo en cada área forman parte de una cultura interna resumida en dos frases que circulan en la operación diaria: “Trabajemos por mejorar lo bien hecho” y “Sin larva no hay paraíso”.
El control de calidad no se detiene en el laboratorio. Se extiende hasta la aclimatación de la larva en la finca, donde personal capacitado acompaña y supervisa el proceso en destino.
Una red de abastecimiento bajo control
El suministro de larvas combina producción interna y alianzas estratégicas. El 70% se produce en el propio laboratorio, mientras que el 30% restante proviene de socios previamente seleccionados. Estos laboratorios asociados son auditados de manera rigurosa, deben contar con certificaciones BAP y el aval de las autoridades ecuatorianas.
Antes de cada cosecha, se analizan muestras con al menos cinco días de anticipación. Solo aquellas que cumplen con los parámetros internos son aceptadas, asegurando que el estándar sanitario se mantenga homogéneo en toda la cadena.
Cuidar el aire, el agua y cada proceso asociado a ellos no es un detalle técnico, sino una decisión que define el rumbo de toda la producción. En la larva se concentra el punto más frágil y, al mismo tiempo, más determinante de la cadena camaronera. Allí se juegan la sanidad, la estabilidad y la confianza del sistema completo. Proteger ese primer eslabón no es una acción aislada, es la base sobre la que se construye todo lo que viene después, porque en larvicultura, el futuro de la producción se decide desde el primer día de vida•
Escanea este código y mira la entrevista completa
Área de desechos que se maneja a través de gestores autorizados
Los laboratorios de larvas: pilares de la resiliencia en la industria camaronera ecuatoriana
Autor:
Yahira Piedrahita
Directora Ejecutiva
Cámara Nacional de Acuacultura
ypiedrahita@cna-ecuador.com
La producción de camarón constituye una de las actividades fundamentales de la economía nacional, destacándose como el principal producto de exportación no petrolero y generando un impacto socioeconómico de gran relevancia. El desarrollo de esta industria, y en particular de su segmento de maduraciones y laboratorios de larvas, constituye un caso ejemplar de adaptación, innovación y sostenibilidad en el sector acuícola global. La transición desde la captura de larvas silvestres de camarón hasta los sistemas de maduración controlada ha estado impulsada tanto por factores de mercado como por la necesidad de enfrentar desafíos sanitarios, ambientales y de sostenibilidad.
Hasta finales de la década de los noventa, la captura de larva silvestre era la principal fuente de semilla para las camaroneras ecuatorianas. Esta práctica, aunque rentable en sus inicios, presentaba serias limitaciones: variabilidad en la oferta, riesgos sanitarios, impacto ambiental y dependencia de factores climáticos como El Niño o La Niña. Aunque hasta fines de esa década, la mayoría de las granjas aún prefería la semilla silvestre por sus supuestos mejores rendimientos, la creciente demanda y la necesidad de asegurar un suministro constante de semilla motivaron la construcción de los primeros laboratorios de producción de postlarvas a principios de la década de 1980.
El laboratorio Semacua, fundado en 1981, fue pionero en la producción comercial de postlarvas en Ecuador, seguido por iniciativas académicas como el laboratorio de la Escuela Superior Politécnica del Litoral (Espol), que además de formar profesionales, suministró semilla a empresas camaroneras. La proliferación de laboratorios permitió una mayor estabilidad en el suministro de larvas;
no obstante, la aparición de enfermedades virales y la presión ambiental fueron los factores que aceleraron la transición hacia sistemas de maduración controlada y el uso de reproductores seleccionados en cautiverio.
Las enfermedades han sido el principal catalizador de la transformación tecnológica y sanitaria en la industria camaronera ecuatoriana. El síndrome de Taura (TSV) en 1994 y el virus de la Mancha Blanca (WSSV) en 1998 provocaron colapsos productivos y pérdidas económicas millonarias. Estos eventos evidenciaron la vulnerabilidad de la industria ante la introducción de patógenos a través de la semilla silvestre y la necesidad de adoptar estrategias de manejo y control sanitario más rigurosas. En respuesta, las empresas comenzaron a implementar programas de selección de reproductores sobrevivientes a las enfermedades que se presentaban en los cultivos, desarrollando líneas genéticas más resistentes y adaptadas a las condiciones locales.
A la par de la crisis sanitaria por Mancha Blanca, en 1998 Ecuador enfrentó una crisis bancaria y financiera severa, con inflación elevada, devaluación del sucre y pérdida de ahorros. Esto llevó a la dolarización oficial en el 2000, sustituyendo nuestra moneda por el dólar estadounidense para frenar la hiperinflación y estabilizar la economía. Durante estos años oscuros, la industria no tuvo acceso a créditos ni financiamiento por parte de la banca privada o pública.
En lugar de rendirse ante la adversidad, los productores demostraron resiliencia y se reinventaron, adoptando nuevas estrategias para garantizar su supervivencia. Los laboratorios de larvas y otros proveedores de insumos hacían trueques o entregaban sus productos con créditos de largo plazo, convirtiéndose en la fuente de financiamiento que permitió que la industria saliera nuevamente a flote. No fue hasta 2003 cuando se observaron los primeros signos de recuperación sostenida de la economía, con un crecimiento del PIB cercano al 3,6%, la inflación controlada en torno al 7% y una mayor estabilidad en el sistema bancario, lo que permitió que la industria volviera a obtener recursos del sistema financiero nacional.
Diagnóstico de los laboratorios de producción de larvas Litopenaeus Vannamei en la provincia del Guayas. Autor: Instituto Nacional de Pesca 2003.
Por otro lado, en 2002 se prohibió de manera indefinida la captura de larvas silvestres de camarón en el Ecuador. Esta medida, que buscaba proteger la biodiversidad marina y reducir el riesgo de introducción de enfermedades, obligó a la industria a depender exclusivamente de la semilla producida en hatcheries, acelerando la adopción de sistemas de maduración controlada y el desarrollo de programas genéticos avanzados.
El mejoramiento genético de los reproductores ha sido uno de los principales motores de la recuperación y el crecimiento sostenido de la industria camaronera ecuatoriana en las últimas dos décadas. Varias empresas han liderado la implementación de estas estrategias, desarrollando líneas genéticas con tasas de crecimiento superiores, mayor supervivencia y resistencia a patógenos. A diferencia de países asiáticos donde predominan sistemas intensivos y el uso de líneas genéticas SPF importadas, Ecuador ha desarrollado líneas propias adaptadas a las condiciones locales y ha priorizado la calidad sobre la cantidad.
Este énfasis en calidad y sostenibilidad ha permitido que el camarón ecuatoriano
se conecte directamente con las nuevas tendencias de consumo saludable en mercados internacionales. En la nueva pirámide alimentaria de Estados Unidos, el camarón ocupa un lugar destacado como proteína magra, baja en grasas saturadas y rica en nutrientes esenciales. Gracias a la innovación en hatcheries y programas genéticos, Ecuador ofrece un producto premium, confiable y uniforme, que responde a la creciente demanda de alimentos funcionales en cadenas de supermercados, restaurantes y plataformas digitales.
La combinación de excelencia productiva, sostenibilidad ambiental y valor nutricional ha posicionado al camarón ecuatoriano como una proteína estrella en el mercado global. Hoy, el país no solo lidera en productividad y calidad, sino que también se alinea con las recomendaciones nutricionales que promueven un mayor consumo de proteínas marinas en dietas equilibradas. Así, la evolución tecnológica de los hatcheries ecuatorianos no solo fortaleció la competitividad de la industria, sino que la conectó con el estilo de vida saludable de millones de consumidores en Estados Unidos y otros mercados clave•
Ecuador alimenta al mundo con su camarón, considerado una superproteína
El camarón ecuatoriano se ha consolidado como uno de los alimentos más valorados a nivel internacional, no solo por su calidad y aporte económico, sino también por sus beneficios nutricionales. En una entrevista concedida a la revista AQUACULTURA, la nutricionista Ana Belén Bermúdez explicó por qué esta proteína ocupa un lugar clave dentro de una alimentación moderna y equilibrada, y cómo su consumo incide positivamente en la salud a lo largo de la vida.
¿Por qué el camarón ecuatoriano es considerado una superproteína?
“El camarón combina un bajo aporte de grasas saturadas con una alta densidad nutricional”, señaló la especialista, “y es una proteína que aporta aminoácidos esenciales, vitaminas, minerales y grasas de buena
Destacó además que esta combinación lo convierte en un alimento funcional, capaz de cubrir requerimientos nutricionales sin un exceso calórico, una característica alineada con las tendencias actuales de alimentación saludable.
Beneficios más allá del desarrollo muscular
Según la nutricionista, el valor del camarón va más allá del fortalecimiento físico. “No solo debemos pensar en crear masa muscular, sino también en prevenir enfermedades cardiovasculares, que son una de las principales causas de muerte a nivel mundial”, explicó. En este sentido, resaltó el contenido de omega-3, específicamente EPA y DHA, que está presente en el camarón y aporta beneficios para la salud del corazón y el sistema nervioso.
“Estos nutrientes también ayudan a la recuperación neuronal y contribuyen a prevenir problemas como la depresión, la demencia o afecciones articulares”, añadió.
"El camarón es ampliamente recomendado después de cirugías o cuando las defensas están bajas".
Ana Belén Bermúdez MSc. Nutrición y Metabolismo
Un alimento para todas las etapas de la vida Bermúdez subrayó que el camarón cumple un rol importante en todas las etapas de la vida, desde la infancia hasta la vejez. “En los niños apoya el crecimiento, el desarrollo muscular y el área neurológica, gracias a su aporte de nutrientes esenciales”, afirmó, “mientras que en los adultos contribuye a la salud digestiva y a la recuperación de masa muscular”.
Para los adultos mayores, su consumo cobra especial relevancia. “Es una proteína de fácil digestión que ayuda a contrarrestar la pérdida natural de masa muscular, conocida como sarcopenia”, puntualizó.
¿Qué lo diferencia de otras proteínas animales?
Al comparar el camarón con otras fuentes de proteína, la nutricionista explicó que su estructura facilita la digestión. “A diferencia de las carnes rojas o procesadas, el camarón
tiene un tejido conectivo más ligero y menos denso, lo que hace que sea mejor tolerado a nivel digestivo”, indicó. Por ello, lo considera una opción adecuada para personas con sensibilidad digestiva, adultos mayores o pacientes en procesos de convalecencia.
Recomendado en procesos de recuperación
Bermúdez también destacó su uso en contextos clínicos: “El camarón es ampliamente recomendado después de cirugías o cuando las defensas están bajas, porque su contenido de selenio ayuda a fortalecer el sistema inmunológico”. No obstante, hizo énfasis en la forma de preparación: “Es importante evitar freírlo o apanarlo en grasas de mala calidad, para que mantenga sus beneficios nutricionales”.
Un alimento estratégico para la salud y el país
Al cierre de la entrevista, la nutricionista resaltó el valor integral del camarón ecuatoriano: “Es un alimento fácil de cocinar, versátil y que se adapta a distintas preparaciones saludables”. Además, añadió que su aporte va más allá de la nutrición individual: se trata de un producto que posiciona al país en los mercados internacionales.
En ese contexto, el camarón ecuatoriano se proyecta no solo como una superproteína dentro de la alimentación moderna, sino también como un símbolo de la capacidad de Ecuador para producir alimentos de alto valor nutricional que llegan a las mesas del mundo•
Escanea este código y mira la entrevista completa
Del valor nutricional al sabor inigualable de una proteína versátil
El camarón es un tema que despierta una profunda pasión en el chef Antonio Hanna, conocido profesionalmente como chef Buman. Fue él quien presentó la Guía Nutricional del Camarón en 2024, consolidándose como una voz autorizada en torno a esta proteína emblemática de la gastronomía ecuatoriana.
Su trayectoria, sin embargo, trasciende el ámbito culinario. En el mundo artístico es reconocido por haber formado parte del grupo musical Los Intrépidos, experiencia que marcó su sensibilidad creativa. Artista integral por naturaleza, Buman ha sabido trasladar esa mirada estética a distintos lenguajes, encontrando en la cocina un espacio donde convergen técnica, identidad y expresión personal.
Por esta razón, el equipo de Revista AQUACULTURA se trasladó hasta su cocina para realizar una entrevista en un ambiente cercano y auténtico, mientras el chef preparaba un exquisito plato a base de camarón y compartía recomendaciones clave para su correcta cocción, resaltando su versatilidad y potencial gastronómico.
De la música a la cocina, ¿en qué momento estos dos mundos se empezaron a fusionar?
Para Buman, la conexión entre su pasado musical y la cocina no es casual, sino parte de una misma forma de entender la creación. Ambas disciplinas se construyen desde la sensibilidad, el ritmo y, sobre todo, el tempo
“Cuando cocino, pienso como músico: busco armonía entre los ingredientes y respeto los tiempos, porque un segundo de más, como una nota mal puesta, puede arruinar el resultado”, asegura.
Esa lógica define su estilo culinario: recetas con pocos elementos, donde nada sobra y todo cumple una función precisa, permitiendo que el producto sea siempre el protagonista.
Todo chef tiene un primer recuerdo con un ingrediente. ¿Cómo fue su primer encuentro con el camarón ecuatoriano en la cocina?
El chef recuerda ese momento con humor y honestidad. No fue una experiencia perfecta, pero sí determinante en su aprendizaje.
“Quedó chicloso”.
Ese error inicial fue clave para entender que el camarón es una proteína que exige técnica y respeto. Con la práctica,
Chef
Buman responde las preguntas de los seguidores de la cuenta de El Mejor Camarón del Mundo, planteadas por Fiorella Echeverría.
Buman aprendió que este producto no admite improvisación y que el punto exacto de cocción marca la diferencia.
Después de probar camarones de distintos países, ¿qué hace que el ecuatoriano sea distinto desde su mirada de chef?
Para él, la respuesta está en el sabor. Tras comparar productos de distintos orígenes, sostiene que el camarón ecuatoriano tiene cualidades únicas.
“Es tan bueno que casi no se necesita adicionarle nada”.
Su dulzor natural, una salinidad equilibrada y una textura firme lo distinguen claramente. Por eso insiste en que cualquier ingrediente debe potenciarlo y no competir con él.
Durante la preparación del plato, la conversación se amplía con la participación de Fiorella Echeverría, integrante del equipo de marketing de la Cámara Nacional de Acuacultura y responsable de las redes sociales de El Mejor Camarón del Mundo Desde su experiencia, traslada a la cocina las preguntas más frecuentes del público recogidas en redes sociales, actuando como un puente entre los consumidores y el chef.
En redes muchos dicen que el camarón es fácil de arruinar. ¿Es realmente así o se trata de un mito?
Buman es claro al responder: no es que el camarón sea un ingrediente difícil, sino mal comprendido. El principal problema, señala, es el miedo a una cocción insuficiente.
“No es difícil, pero sí mal entendido”.
Explica que muchas personas lo cocinan en exceso por precaución, cuando en realidad eso afecta directamente su textura y sabor.
Entonces, ¿cuál es el punto perfecto de cocción del camarón?
Para el chef, este es el momento más importante de toda la preparación. Asegura que la mayoría de errores no tienen que ver con el condimento, sino con pasarse de cocción. El camarón es una proteína sensible que cambia en cuestión de segundos.“El camarón está listo cuando pasa de gris a rosado y la carne queda firme, pero jugosa”. En agua hirviendo, el tiempo es exacto. “Tres minutos por reloj y no más”, explica, advirtiendo que prolongar ese tiempo lo vuelve chicloso. En sartén o a la plancha, la clave es el sellado rápido. “Menos de un minuto por lado es suficiente”, señala, siempre con el fuego alto y el camarón bien seco antes de entrar al calor.
Para Buman, no se trata de intuición, sino de atención y precisión. Retirarlo a tiempo es una forma de respeto al producto.
Otra duda recurrente en casa: ¿siempre hay que marinar el camarón?
Ante esta pregunta frecuente, el chef aclara que no es necesario. El camarón ecuatoriano tiene sabor propio y no requiere excesos. “Mientras menos, mejor”.
Para él, basta sazonar con sal, pimienta y, si se desea, un toque sutil de paprika o ají. Saturarlo de condimentos le quita protagonismo.
En una cocina más consciente, ¿qué tanto del camarón se puede aprovechar?
Buman destaca que el camarón debe utilizarse de manera integral.
Cabezas y cáscaras son ideales para la elaboración de fondos, caldos, aceites y sales.
“El secreto de una buena cocina está en los buenos fondos”.
Desde su perspectiva, el camarón ofrece un enorme potencial más allá del plato principal.
Desde su experiencia, ¿qué convierte al camarón en una proteína tan versátil?
El chef lo resume con claridad. “Se adapta a todo”. Puede ser protagonista de un plato cotidiano o de una preparación especial, integrarse a pastas, parrillas o recetas más elaboradas sin perder identidad.
Para cerrar, el camarón ecuatoriano tuviera que definirse en una sola idea, ¿cuál sería?
Buman concluye con una reflexión que conecta gastronomía e identidad nacional.
“Tenemos un tesoro y no siempre lo valoramos”.
Para él, el camarón ecuatoriano es un lujo reconocido en el mundo, pero muchas veces subestimado dentro del país. Su invitación final es clara: perder el miedo, cocinarlo con respeto y sentirse orgullosos de un producto que lleva el nombre de Ecuador a lo más alto. Entre arte, técnica y la interacción con su público, el camarón reafirma su lugar como una proteína versátil, noble y excepcional, capaz de brillar cuando se le concede el espacio que merece•
Escanea este código y mira la entrevista completa
LPamela Nath Directora
a acuicultura no suele aparecer en los sueños de infancia. No es una profesión que se imagine jugando ni una palabra frecuente en las conversaciones escolares. Y, sin embargo, para muchas personas termina siendo un lugar de destino. Un espacio donde se abren oportunidades y se construyen trayectorias profesionales.
La industria camaronera en Ecuador ofrece justamente ese tipo de oportunidades. Es uno de los principales generadores de empleo del país y una fuente de trabajo formal en zonas rurales que no cuentan con vocación agrícola ni alternativas productivas. Cerca de 300 mil plazas directas e indirectas dependen de esta actividad, y más de 160 mil corresponden a empleo directo, según datos de la Cámara Nacional de Acuacultura (CNA).
Dentro de esa fuerza laboral, más de 50 mil mujeres trabajan hoy de forma directa en el sector. Para muchas de ellas (en especial jefas de hogar), este empleo representa autonomía económica, estabilidad y la posibilidad de sostener a sus familias. En la práctica, el acceso a un trabajo formal y estable es uno de los pilares más efectivos para avanzar hacia la igualdad de género.
La industria camaronera también ha ampliado los espacios de desarrollo profesional para las mujeres. Su presencia ya no se concentra únicamente en tareas operativas, sino que se extiende a funciones técnicas, administrativas y de liderazgo. La Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) destaca que la participación plena de las mujeres en todos los eslabones de la cadena productiva acuícola fortalece al sector y contribuye a su sostenibilidad.
A través de este reportaje mostramos cómo en Ecuador, varias mujeres han reconocido ese potencial y han apostado por una industria que aún se sigue construyendo. Desde distintos caminos, han puesto sus capacidades al servicio del desarrollo del sector y han trazado trayectorias profesionales que les permiten ejercer liderazgo, tomar decisiones y proyectar futuro. El suyo y el del sector.
Alos 17 años, Annabell Palacios tenía claro que quería estudiar medicina. De niña jugaba a curar patitos y pollitos en el entorno rural en el que creció, ubicado en la provincia de El Oro, en Ecuador. “Jugaba a cuidar”, cuenta, como si esa vocación hubiese estado ahí desde siempre.
Pero ese camino no fue posible. Las condiciones económicas pesaron y también una forma de pensar muy extendida en ese momento: la idea de que, como mujer, pronto se casaría y formaría su hogar, y que estudiar no era una prioridad. Cuando pidió apoyo para continuar, la respuesta fue “no”.
“Era un rol bastante equivocado del potencial que tienen las mujeres”, reflexiona. Y reconoce que ese límite fue, paradójicamente, el que le hizo tener “todavía más ganas de salir adelante”.
Buscó oportunidades en la Universidad Técnica de Machala. Analizó todas las carreras disponibles. Se preguntó cuál le permitiría manejar su tiempo, trabajar al aire libre, ser productiva y, sobre todo, crecer en una provincia marcada por la actividad agrícola.
Así, descubrió la acuicultura de camarón, un sector nuevo que empezaba a ganar espacio. “Vi que tenía potencial, que había necesidades profesionales y sentí que yo lo podía hacer bien”, recuerda.
Con el tiempo entendió que no había renunciado a su vocación inicial. En la acuicultura encontró muchas de las razones que la habían
Annabell Palacios, CEO de Agrimingold
llevado a pensar en la medicina: trabajo constante, decisiones críticas y cuidado de seres vivos. También encontró algo más: libertad.
Cuando se le pregunta por su cargo y la frase queda en suspenso, “¿usted es CEO de…?”, Annabell se ríe antes de completar la idea: “Soy CEO de mi vida, primero que nada”. Luego aclara: es CEO de Agrimingold, una empresa familiar productora de camarón, y también de la comercializadora Marea Rosa; además, es una de las fundadoras de la Organización Mundial de Mujeres en la Acuicultura (OMMA).
Hoy vive la producción en el día a día, y tiene claro que las 30 personas que trabajan junto a ella en Agrimingold pasan más tiempo en la finca que en sus propias casas. Por eso establece un principio no negociable: si allí ella duerme en condiciones dignas, su equipo también debe hacerlo. “Se trata de una igualdad que creo que es justa”, afirma.
Su carácter se refleja también en sus decisiones productivas. Cuenta que trabajan desde la prevención, que capacitan constantemente a su personal, y que desde hace más de una década no usan antibióticos. “Usarlos sería retroceder”, explica, “porque podemos recuperar una piscina, pero no la credibilidad”.
Desde ese lugar, Annabell percibe que el camino para las mujeres en la industria ya está abierto, aunque reconoce que cuando ella empezó era más difícil. Cree que muchas veces los límites están más en el miedo que en las puertas cerradas. Señala que ya no se da solo por inclusión, sino por reconocer que la industria necesita lo que las mujeres aportan: empatía, comunicación, organización y liderazgo.
Esa misma decisión, apostar por la acuicultura y quedarse, aparece también en otras trayectorias del sector.
Cuando María Claudia Baquero terminó sus estudios de Biología Marina en la Universidad Atlántica de Florida, en Estados Unidos, sabía que quería dedicarse a la “piscicultura o la camaronicultura”. Le recomendaron ir al sur de Colombia a trabajar con peces. Tenía sentido, pues se trataba de su país de
origen. Pero no sucedió. “La intuición me dijo que era mejor trabajar en la producción de camarón”, recuerda. Así, decidió migrar a Ecuador.
“Fue la mejor decisión de mi vida”, afirma.
Hoy, María Claudia es jefa de área en Naturisa, donde coordina los procesos productivos de la finca y lidera a un equipo de alrededor de 40 personas. En producción, “cualquier equivocación de análisis puede generar un error irreversible”. Sobre todo, en un entorno que cambia todo el tiempo y exige adaptabilidad. Trabaja con animales vivos y con decisiones que afectan a toda una cadena de procesos.
Por eso, antes de decidir, necesita entender el estado de ese sistema: cómo amanecieron las piscinas, si hubo desajustes en oxígeno o temperatura durante la noche, y cómo respondió el camarón a la alimentación. Con esa información define prioridades y ajusta la operación para evitar errores que se arrastran en cadena.
María Claudia sabe que liderar en producción implica operar en función de variables como el clima, el cambio en el manejo y la tecnología. Por eso valora habilidades como la adaptabilidad, la proactividad y la capacidad de resolver problemas. En el
trabajo con el equipo, prioriza la colaboración, la comunicación clara y una buena relación. “Para mí es muy importante la empatía y la preocupación por las emociones de cada uno de los integrantes”, señala.
De niña, María Claudia se imaginaba siendo bióloga marina y liderando equipos. Al elegir la acuicultura de camarón en Ecuador, encontró un lugar donde esa visión tomó forma en el trabajo diario.
Y sostener esa visión en el tiempo también exige ordenar los procesos y criterios que rodean a la producción.
María Claudia Baquero, jefa de área en Naturisa
SOSTENIBILIDAD
Kenya Guerra siempre tuvo claro que quería trabajar en producción sostenible. Durante varios años lo hizo desde la industria bananera, enfocada en certificaciones, sostenibilidad y gestión ambiental. El camarón apareció después. “No solo por afinidad personal”, explica, “sino porque entendía su importancia para el Ecuador como principal producto de exportación y motor económico del país”.
Cuando se presentó la oportunidad de incorporarse a Promarisco, una compañía que se dedica al cultivo y procesado del camarón, decidió dar el paso. “Me encontré un sector muy dinámico, técnicamente desafiante y con un enorme potencial de mejora continua”, explica. Eso fue lo que la hizo quedarse. “Descubrí el impacto que podía tener la acuicultura desde mi formación técnica y mis valores profesionales”, confiesa.
Hoy, como jefa de Sostenibilidad y Control de Procesos, entiende que el alcance de su trabajo es transversal. Sus decisiones inciden en distintas áreas de la operación, desde el cumplimiento ambiental y la bioseguridad hasta la trazabilidad y la preparación frente a auditorías.
“La sostenibilidad requiere equilibrio entre productividad y cuidado ambiental, entre eficiencia y bienestar laboral, entre cumplir estándares y generar cultura”, cuenta. Liderar desde ahí implica escuchar, adaptarse y generar compromiso.
“Los mejores resultados se logran cuando los equipos comprenden el por qué de cada práctica”, afirma, “y cuando la educación interna pesa tanto como los indicadores”. Desde su posición, Kenya observa que cada vez más profesionales con esa mirada (muchas de ellas mujeres) están ocupando espacios técnicos y de decisión dentro de la industria.
Cuando piensa en su futuro, se imagina creciendo dentro de áreas que integren sostenibilidad con estrategia corporativa, innovación o gobernanza ambiental. Le interesa continuar contribuyendo y generar transformaciones reales a escala en el sector. Eligió la acuicultura pensando en el largo plazo: “es un sector vivo, que evoluciona rápido, y eso me motiva cada día”.
En esta industria hay decisiones que no se explican solo mirando desde adentro, sino desde lo que ocurre fuera.
Ana María Costa trabaja en ese punto. Desde su rol como gerente de Sistemas de Gestión en la productora y exportadora de camarón Sociedad Nacional de Galápagos (Songa), está a cargo de responder a las exigencias de los mercados internacionales, esos requisitos que pueden definir si una exportación avanza o se detiene.
Es ingeniera en alimentos, con especialización en ciencia de alimentos, y cuenta con más de 18 años de experiencia en calidad, inocuidad y producción dentro de la industria alimentaria. En Songa lleva alrededor de 8 años. A lo largo de su carrera ha participado en más de 300 auditorías, tanto de autoridades competentes como de certificaciones.
Su trabajo se centra en asegurar la calidad del producto: controles, trazabilidad, inocuidad, manejo de certificaciones en plantas, fincas camaroneras y laboratorios de larva, y habilitaciones que deben cumplirse para poder exportar. Desde su experiencia, observa que los mercados cambian y que las exigencias se actualizan de forma constante. “Con base en requerimientos de clientes y de mercados de destino, nos ha tocado modificar las formas con las que trabajamos tradicionalmente”, explica. Por eso, su trabajo implica revisar y ajustar procesos
de manera continua. “La industria no puede conformarse, debe ir evolucionando”, dice.
Lejos de ver esa exigencia como una carga, Ana María encuentra ahí el valor de su profesión. La calidad obliga a mantenerse informada, a aprender de forma continua y a no caer en la monotonía. Para ella, esa capacidad de adaptación es la que permite que el camarón ecuatoriano siga compitiendo en mercados cada vez más exigentes.
Responder a esas exigencias también implica tomar decisiones de crecimiento.
Verónica Dahik aprendió temprano a no dejar que el miedo decida por ella. “¿Por qué hay que tener miedo?”, pregunta con naturalidad y se responde, “lo peor que puede pasar es que te digan que no”. Esa idea ha guiado su recorrido en la industria camaronera durante los últimos 18 años.
Llegó al sector cuando tenía 23. Retomó un proyecto familiar que aún no se había puesto en marcha: una infraestructura ubicada en Playas, en la provincia del Guayas, concebida inicialmente para producir carnada. Lo analizó. Miró el entorno, el mercado y las posibilidades reales. Playas es un puerto pesquero y el camarón era un producto con demanda y posibilidades de comercialización. Cambió el enfoque y comenzó a trabajar con camarón de mar.
Así nació Natluk, una empresa empacadora y exportadora de camarón, dedicada tanto
Kenya Guerra, jefa de Sostenibilidad y Control de Procesos en Promarisco
Ana María Costa, gerente de Sistemas de Gestión en Songa
al camarón de mar como al de acuicultura, que hoy procesa y comercializa producto ecuatoriano para mercados internacionales.
“No temer a los retos y ser resilientes ha sido una de las razones por las que hemos
ido creciendo”, cuenta. Otra razón ha sido su foco constante en dos conceptos: innovación y mejora continua. “Hay que tener tiempo para reflexionar dónde estás y analizar cuáles son tus siguientes pasos”, comparte. Por ejemplo, ella se plantea crecer al menos un 20% en productividad cada dos años. Hoy, como CEO de Natluk, su rol implica tomar decisiones, liderar equipos y sostener una operación que no se detiene. “Esto es 24/7”, dice.
Hay una motivación que atraviesa su forma de mirar el negocio. Aunque es de Guayaquil creció en Playas, donde recuerda que no había trabajos estables. Hoy ve empleo formal en la zona y sabe que su empresa forma parte de ese cambio. “Mientras una parte del trabajo me genera actividad, esto me llena de propósito”, expresa.
Cuando piensa en el futuro de la industria, Verónica mira más allá del volumen.
SOSTENIBILIDAD
“Quiero ver cómo Ecuador crece no solo en exportaciones, sino en innovación”, dice. Cree que el liderazgo del país como principal productor mundial de camarón abre una oportunidad: hacer mejor las cosas y agregar valor, innovando en los procesos, en los estándares y en la forma en que el producto se presenta y llega al mercado.
Este año, Natluk abrirá nuevas instalaciones productivas que le permitirán aumentar su productividad entre un 30 y 40%. Para Verónica, ese paso es parte del mismo proceso que la ha traído hasta aquí.
Apostar, ajustar y crecer. En una industria que todavía se sigue construyendo, las voces de estas mujeres empujan ese proceso hacia adelante, sin miedo al “no”•
Verónica Dahik, CEO Natluk
Se reformula el combate a la delincuencia en el Ecuador
a implementación de la Estrategia Operacional 3D marca un punto de inflexión en la política de seguridad del Ecuador y se consolida como un pilar clave para la protección de sectores estratégicos como la industria camaronera. En un escenario complejo, donde la delincuencia organizada busca presionar economías formales, el país avanza hacia un modelo de seguridad más articulado, preventivo y sostenido, que combina acción policial, tecnología de punta y coordinación con el
Desde esta nueva lógica, la industria camaronera ha asumido un rol activo y corresponsable. El sector ha destinado una inversión cercana a los 100 millones de dólares anuales en seguridad, orientada a la implementación de sistemas avanzados de videovigilancia, monitoreo permanente,
suma la participación activa de la Cámara Nacional de Acuacultura (CNA) en mesas de trabajo de seguridad junto al sector productivo, un espacio que garantiza comunicación directa, actualización permanente de información y la posibilidad de reportar de inmediato cualquier incidencia en zonas camaroneras a nivel nacional. Este esquema ha permitido pasar de una lógica reactiva a una preventiva, donde los riesgos se identifican y gestionan con mayor anticipación.
El trabajo conjunto se desarrolla en un contexto donde persisten ciertos delitos que siguen aquejando a los productores, principalmente en zonas de la Costa. Entre ellos se registran intentos de extorsión, robos a instalaciones, sustracción de motores, equipos e insumos, así como asaltos esporádicos en rutas utilizadas para el transporte de personal y producto. No obstante, tanto autoridades como gremios coinciden en que estos hechos no se han generalizado ni escalado, y se mantienen contenidos gracias a la acción preventiva, el control territorial y la respuesta coordinada.
La contención de estos riesgos forma parte de un proceso más amplio de transformación de la política de seguridad. Ecuador avanza hacia nuevas formas de enfrentar la delincuencia organizada, dejando atrás respuestas fragmentadas para consolidar un enfoque institucional, planificado y sostenido en el tiempo. En ese marco, la Policía Nacional, a través de sus unidades especializadas, ha fortalecido su presencia operativa y su capacidad de coordinación interinstitucional.
mantenimiento de turnos rotativos de guardias privados que operan de manera continua en zonas de alta sensibilidad.
Este esfuerzo se complementa con acuerdos de cooperación con el sistema ECU 911, lo que permite un control más eficiente de la vigilancia en áreas identificadas como de alto riesgo.
A esta articulación se
De igual forma, el diseño de los operativos responde a objetivos claramente definidos. “Perseo” se enfoca en intervenciones policiales intensivas en sectores con alta incidencia delictiva, reforzando el control territorial.
“Pandora” actúa sobre espacios donde se detectan economías ilícitas, como centros nocturnos o puntos informales utilizados para actividades criminales. “Apolo”, por su parte, apunta a la desarticulación de estructuras de alta peligrosidad, mediante operaciones selectivas basadas en inteligencia previa.
Este esquema se inscribe dentro de la Estrategia Operacional 3D, denominada así porque está orientada a debilitar, desorganizar y desarticular al crimen organizado. Uno de sus componentes más relevantes es
la incorporación de tecnología aplicada a la seguridad, como el código QR de proximidad comunitaria, que fortalece la denuncia oportuna y permite una respuesta más rápida y focalizada. Para el sector camaronero, este componente ha sido clave. La posibilidad de reportar de forma inmediata situaciones sospechosas ha permitido anticipar escenarios de riesgo, orientar patrullajes y ejecutar intervenciones oportunas, convirtiendo la información en un insumo estratégico para la labor de inteligencia.
Las autoridades destacan que uno de los principales logros de esta estrategia ha sido evitar una escalada de violencia en zonas productivas. Si bien los delitos continúan presentándose de manera puntual, no han logrado consolidarse como prácticas sistemáticas.
La presencia policial, el patrullaje coordinado y el seguimiento permanente han generado un entorno menos favorable para la acción delictiva.
Desde el sector privado, la respuesta ha sido igualmente estratégica. La industria camaronera ha entendido que la seguridad no se limita a la reacción, sino que requiere prevención, planificación y corresponsabilidad, protegiendo así miles de empleos directos e indirectos, y garantizando la continuidad de una actividad clave para la economía nacional.
El Gobierno Nacional ha reiterado que la lucha contra la delincuencia organizada es un proceso de mediano y largo plazo, que exige continuidad y evaluación permanente.
La experiencia reciente demuestra que la coordinación institucional, el uso de tecnología y la cooperación con sectores estratégicos constituyen hoy una de las herramientas más eficaces para enfrentar al crimen organizado.
Aunque el desafío de la delincuencia organizada sigue presente, los avances logrados a través de la Estrategia Operacional 3D demuestran que es posible contener el delito, reducir su impacto y limitar su capacidad de expansión, especialmente cuando existe coordinación efectiva entre el Estado y los sectores productivos.
El camino no está concluido, pero sí trazado. La combinación de planificación estratégica, uso inteligente de la tecnología, presencia territorial y corresponsabilidad privada ha permitido construir un entorno más controlado y previsible, donde el crimen ya no opera con la misma libertad ni define las condiciones del desarrollo económico●
ARTÍCULOS TÉCNICOS ÍNDICE
Edición 169 - Febrero 2026
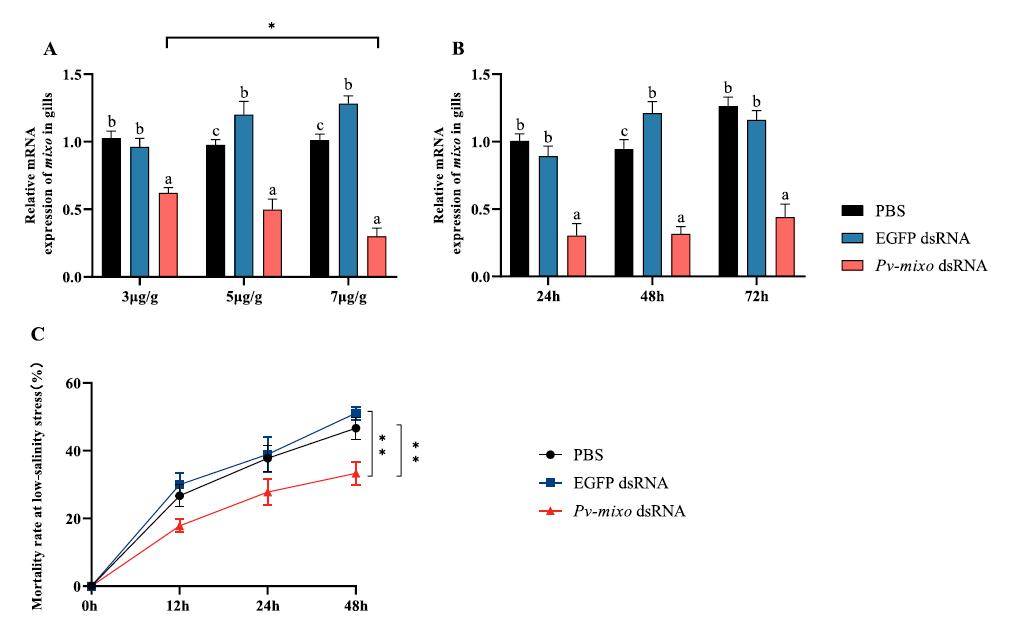
Enfermedad de la Postlarva Translúcida: una infección mortal por Vibrio que afecta al camarón de cultivo
Desbloqueo del potencial de la microbiota: el papel del cobre orgánico en la mejora del cultivo saludable del camarón blanco (Penaeus vannamei)
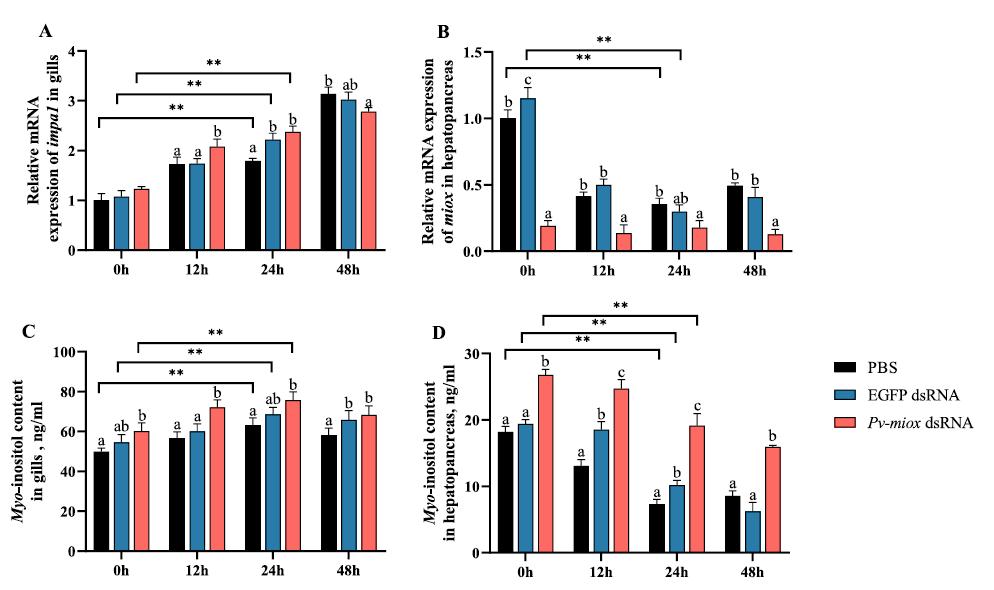
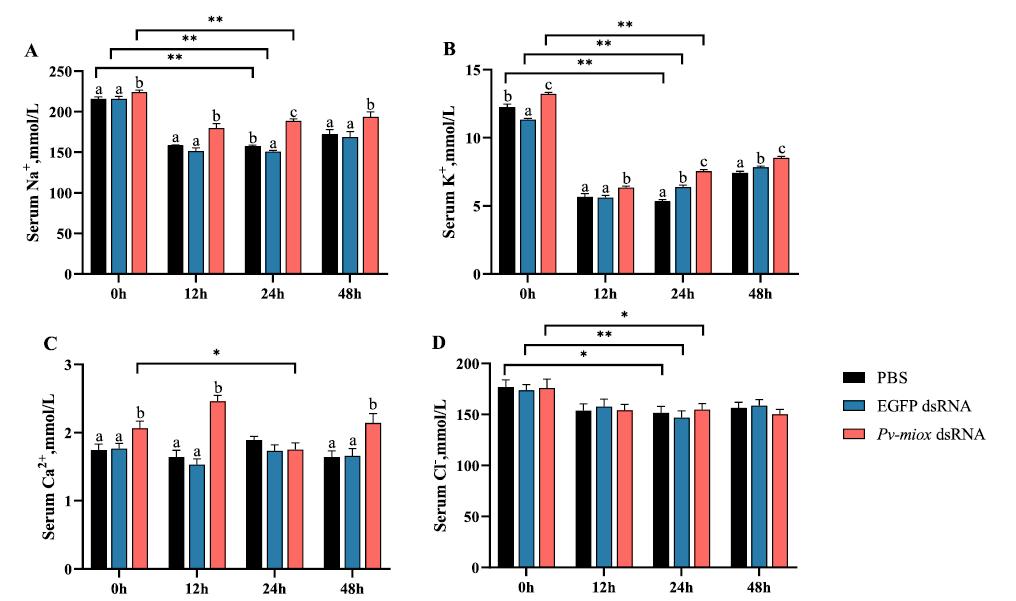
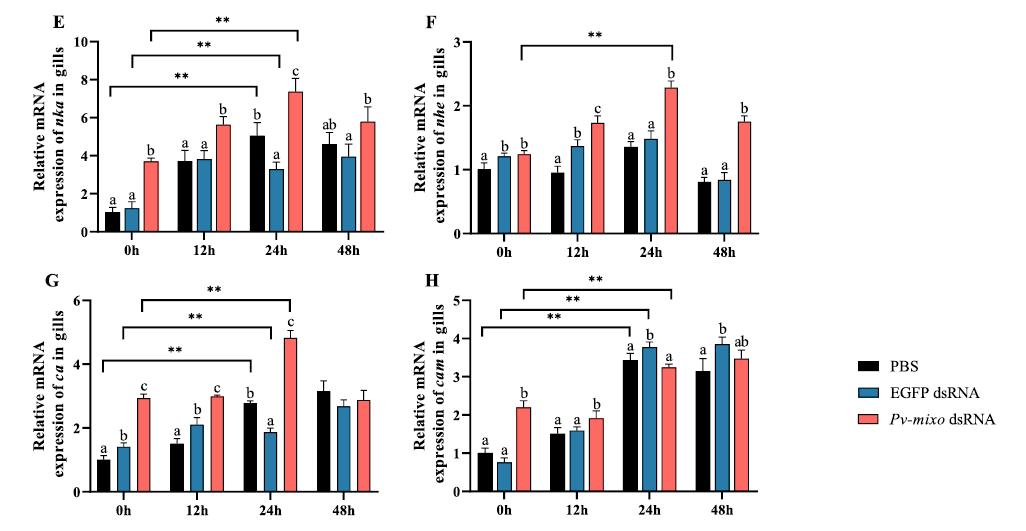
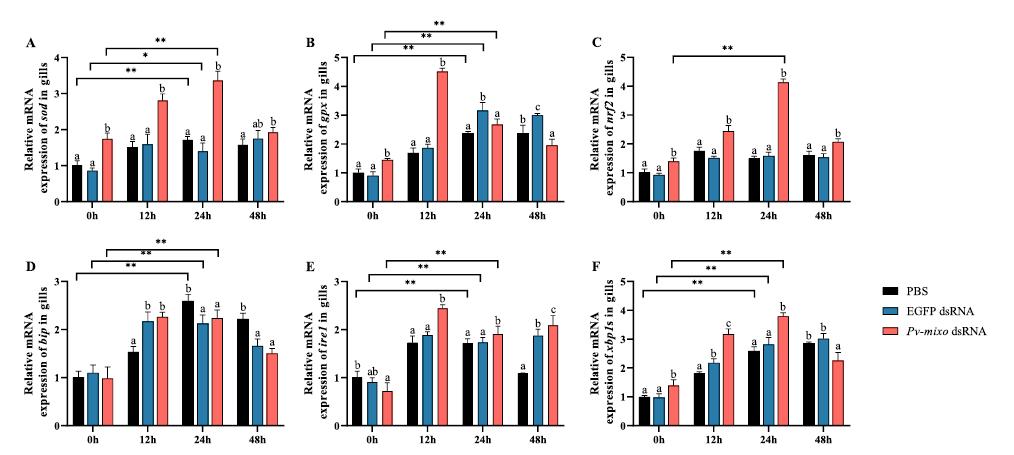
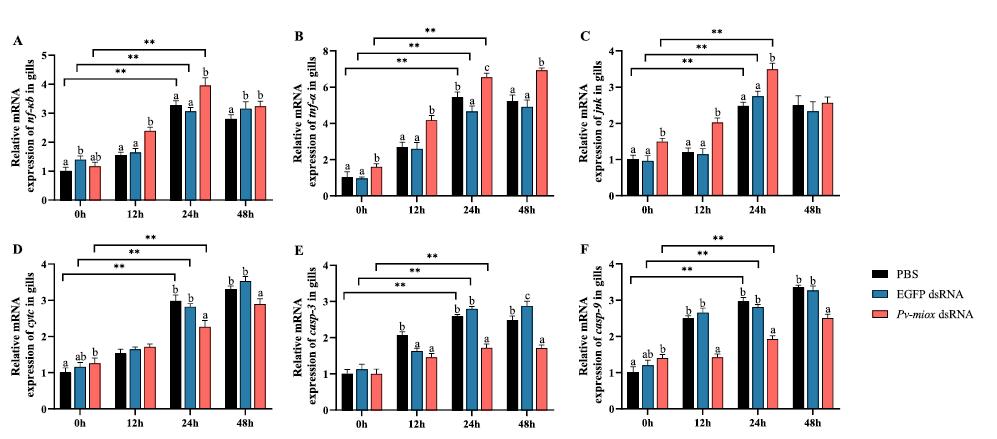
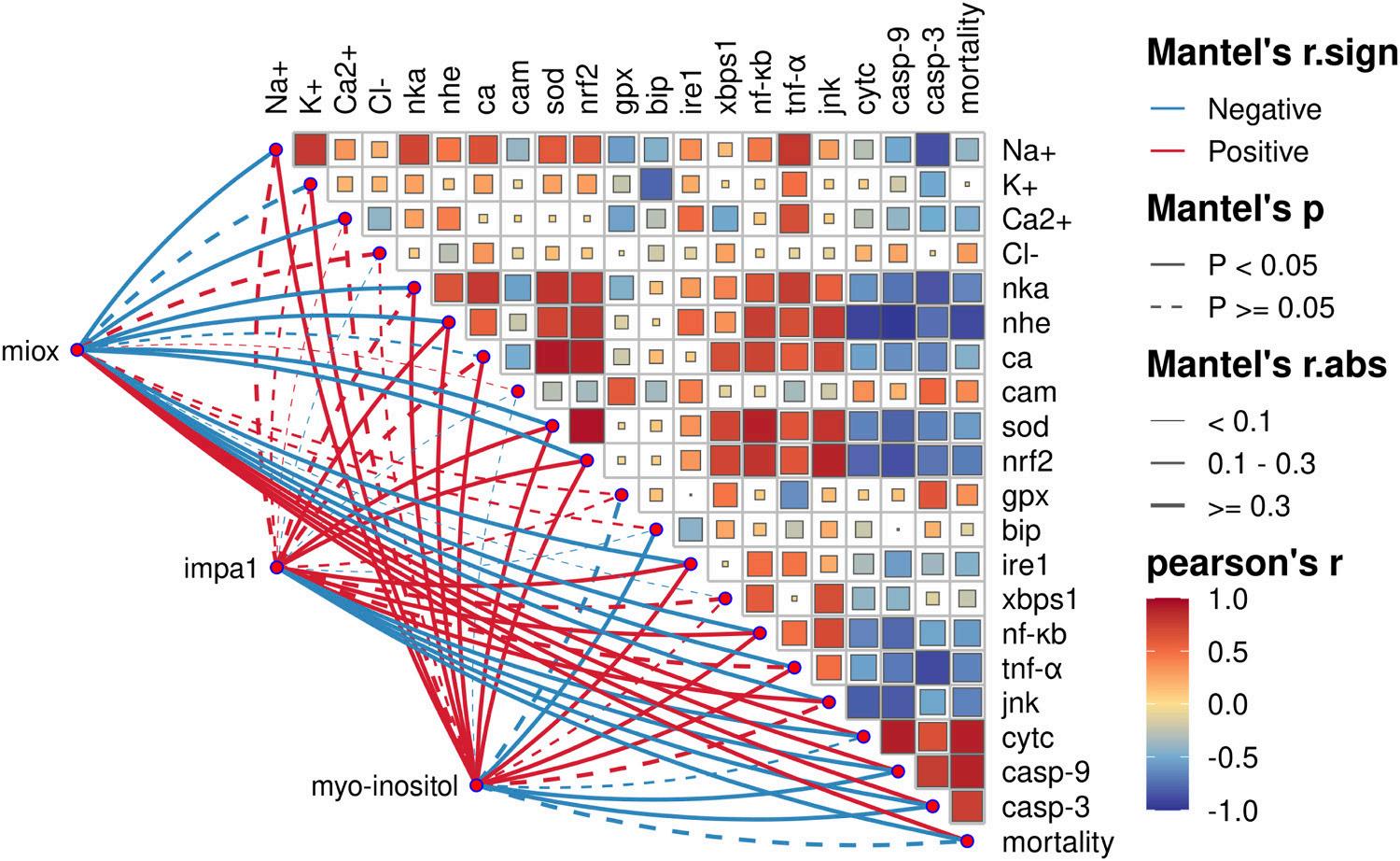
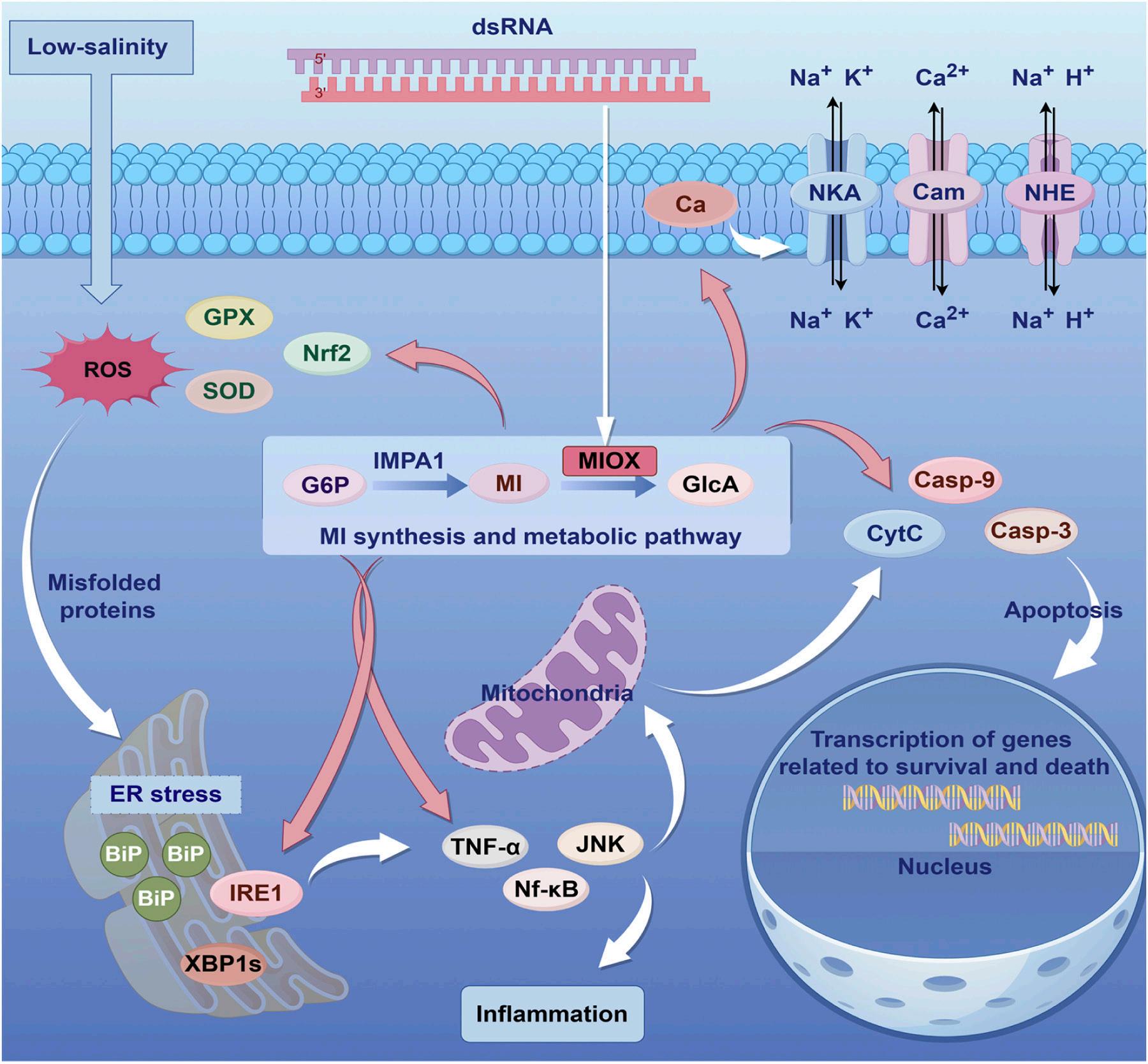
Mioinositol oxigenasa: un actor clave en la lucha del camarón blanco del Pacífico (Penaeus vannamei) contra el estrés por baja salinidad
Efectos de un co-cultivo inactivado por calor de Lactobacillus spp. (postbiótico) sobre el rendimiento de crecimiento y la tolerancia al estrés ambiental en postlarvas de camarón blanco (Penaeus vannamei): síntesis de cuatro ensayos independientes
Técnicas recomendadas para la biorremediación del estuario del Guayas y su área de influencia el perfil costero–marino de la provincia de Santa Elena
Enfermedad de la Postlarva Translúcida: una infección mortal por Vibrio que afecta al camarón de cultivo
Las pruebas moleculares son esenciales para la identificación precisa de la causa específica, junto con estrictas medidas de bioseguridad.
Figura 1: Las postlarvas infectadas con TPD en los estadios 2 a 4 presentan un intestino vacío y un hepatopáncreas incoloro y translúcido. La mortalidad suele ocurrir a las pocas horas de la infección. Foto: Xu Tao
Comprendiendo las infecciones por Vibrio en la producción de camarón
Las infecciones asociadas con especies de Vibrio plantean desafíos significativos en las operaciones comerciales del camarón. Si no se controlan, estas bacterias pueden establecer poblaciones en el agua, sedimentos o biofilms dentro de los sistemas de cultivo, lo que provoca infecciones y altas tasas de mortalidad.
En los laboratorios de camarón, las especies de Vibrio pueden infiltrarse o proliferar a través de múltiples vías. Estas incluyen su introducción a través de reproductores, nauplios de camarón infectados, contaminación a través de fuentes de agua, transmisión por microalgas, alimento vivo o tuberías de agua y aire. Además, pueden transportarse en el equipo del personal, en la piel o dispersarse a través de aerosoles (Shinn et al., subm.).
¿Qué
es la enfermedad de las Postlarvas Translúcidas?
La enfermedad de las Postlarvas Translúcidas (TPD), también conocida como enfermedad de Vibrio Altamente Letal (HLVD), es una afección grave que ha afectado la producción de postlarvas de camarón en China y Vietnam desde 2020.
La enfermedad es causada principalmente por una cepa de Vibrio parahaemolyticus, aunque también se ha implicado un Baishivirus en algunos casos. La cepa de V. parahaemolyticus asociada con la TPD produce una toxina que altera el hepatopáncreas, lo que afecta la nutrición y provoca una rápida mortalidad, especialmente en camarones más pequeños.
Visualmente, las postlarvas afectadas (PL), particularmente en los estadios 2-4, presentan síntomas distintivos, como un intestino vacío y un hepatopáncreas incoloro y translúcido, lo que provoca una disminución de la actividad y movimientos lentos. La mortalidad ocurre rápidamente, generalmente en unas pocas horas después de la infección, con tasas que alcanzan hasta el 80-100% en 2448 horas, y que a menudo ocurren entre 3 y 5 días después de la repoblación.
Dado que otros patógenos pueden causar síntomas visuales similares en el camarón, un diagnóstico preciso requiere pruebas adecuadas en lugar de basarse únicamente en los síntomas visibles.
El papel de Vibrio parahaemolyticus en la TPD
La mayoría de los casos de TPD son causados por cepas de V. parahaemolyticus portadoras del gen de la aerolisina, que produce una toxina que causa daño celular en el hepatopáncreas y causa la muerte. Otra enfermedad del camarón, la Necrosis Hepatopancreática Aguda (AHPND), también es causada por V. parahaemolyticus, pero estas cepas portan un gen de toxina diferente. Si bien ambos genes de toxina causan daños similares al hepatopáncreas y conducen a resultados comparables, las pruebas moleculares son esenciales para la identificación precisa de la causa específica.
¿Cómo detectar y distinguir TPD y AHPND?
Las inspecciones visuales por sí solas no son suficientes para
diagnosticar TPD o AHPND. Un diagnóstico preciso requiere pruebas de laboratorio. Para ello, se debe recolectar una muestra específica de PL (es decir, aquellas que presentan características pálidas y moribundas, aproximadamente 30-50 individuos). Enjuagar las PL con agua destilada estéril y luego fijarlos en etanol de grado molecular al 95-99%.
Las pruebas de laboratorio pueden identificar los diferentes genes productores de toxinas producidos por las especies de Vibrio. Simultáneamente, se debe solicitar que las muestras se analicen para detectar la presencia de genes plasmídicos pirAB que producen las toxinas responsables de la AHPND, utilizando el método de PCR anidada AP4 desarrollada por Dangtip et al. (2015) y para TPD utilizando ptimers para el gen ldh, que produce la toxina hemolisina termolábil (Vicente et al., 2020; Zou et al., 2020). Si ambas pruebas son negativas, se debe solicitar que las muestras se analicen para el Baishivirus, utilizando primers según lo especificado por Xu et al. (2023).
La evaluación paralela de los resultados microbiológicos es esencial. Si los tres resultados de la prueba PCR son negativos, hay que buscar otros posibles patógenos bacterianos que podrían resultar en PL de apariencia translúcida.
¿Por qué realizar la prueba?
Las pruebas regulares son cruciales para identificar infecciones en etapa temprana y prevenir su establecimiento y propagación, además de aumentar la conciencia sobre los riesgos locales de infección. Las pruebas pueden ofrecer información valiosa sobre la dinámica de la enfermedad y las posibles rutas de introducción. Facilitan intervenciones oportunas y permiten la revisión de los protocolos de bioseguridad para reducir la probabilidad de futuras introducciones y brotes, minimizando así las pérdidas económicas.
“Los Vibrios representan una grave amenaza para la producción de camarón, pero medidas estrictas de bioseguridad y la vigilancia regular pueden gestionar eficazmente estos riesgos”.
Manejo de brotes de TPD
Si se detecta TPD, tome medidas inmediatas:
• Aislar los camarones infectados y poner en cuarentena las áreas afectadas.
• Realizar pruebas para confirmar la enfermedad.
• Evaluar el riesgo para otros lotes de camarones y para las operaciones generales de la camaronera.
• Eliminar las poblaciones infectadas para evitar una mayor propagación.
• Reforzar las medidas de bioseguridad, incluyendo el monitoreo de visitantes y la desinfección de equipos y sistemas de agua.
• Aumentar la vigilancia para monitorear la situación y prevenir futuros brotes.
Otras
infecciones
por Vibrio
que afectan a los camarones
Además de la TPD, otras infecciones por Vibrio también pueden causar que los camarones se vean translúcidos. Por ejemplo, algunas cepas de V. parahaemolyticus que portan un gen de toxina diferente son responsables del AHPND, también conocido como síndrome de Mortalidad Temprana, que puede provocar una mortalidad repentina y grave. Además, otras especies de Vibrio, como V. alginolyticus y V. harveyi, pueden causar infecciones sistémicas que conducen a la Necrosis Hepatopancreática Séptica (SHPN).
Necesidad de vigilancia y bioseguridad estrictas para Vibrio
La rápida y grave aparición de mortalidades inducidas por Vibrio en criaderos de camarones peneidos, que pasan de un estado de aparente salud a moribundos y muertos en cuestión de horas, resalta la necesidad crítica de medidas de bioseguridad rigurosas y protocolos de vigilancia rigurosos.
Reconocer los riesgos asociados a las infecciones por Vibrio es fundamental desde el punto de vista de la bioseguridad, lo que requiere medidas proactivas para prevenir y mitigar posibles brotes. Establecer procedimientos sólidos de control y gestión es esencial para gestionar eficazmente estos riesgos. La vigilancia se convierte en una práctica crucial para la detección temprana y la contención de infecciones, garantizando una intervención rápida cuando sea necesario.
Mejores prácticas de bioseguridad en criaderos de camarones:
1. Desinfectar el agua y el equipo con ozono, luz ultravioleta o hipoclorito.
2. Realice una limpieza exhaustiva y regular de todo el sistema de producción, incluyendo tuberías, líneas de aire y sistemas de suministro de aire, para eliminar biopelículas y superficies donde Vibrio pueda establecerse.
3. Utilice sistemas de agua separados y bioseguros para minimizar los riesgos de contaminación.
4. Añada probióticos al agua para mejorar la salud de los camarones y reducir las bacterias dañinas.
5. Aísle los reproductores en condiciones limpias y proporcione dietas bioseguras para mantener su salud.
Manejo de Vibrio
en cada etapa de la producción de camarones
Las infecciones por Vibrio pueden ocurrir en cualquier etapa del cultivo de camarón, desde los reproductores hasta los estanques de engorde. Implementar una desinfección adecuada, mantener una higiene estricta y añadir probióticos al alimento y al agua son esenciales para reducir los riesgos de infección.
• Criaderos: garantice sistemas de agua bioseguros y realice limpiezas regulares de las instalaciones.
• Alimento vivo: adquiera alimentación viva de proveedores bioseguros para prevenir la introducción de Vibrio
• Preparación de los estanques: desinfecte los estanques, utilice agua limpia y evite transferir agua contaminada.
• Estanques de engorde: monitoree la densidad de población y gestione los desechos eficazmente para minimizar el riesgo de brotes de Vibrio.
Conclusiones
Las bacterias Vibrio representan una grave amenaza para la producción de camarón, pero las estrictas medidas de bioseguridad y la vigilancia regular pueden controlar eficazmente estos riesgos. La detección temprana es crucial para minimizar el impacto de las enfermedades relacionadas con Vibrio, mejorar la supervivencia del camarón y garantizar la sostenibilidad de las operaciones de cultivo•
Desbloqueo del potencial de la microbiota: el papel del cobre orgánico en la mejora del cultivo saludable del camarón blanco (
La secuenciación de la microbiota se ha convertido en una herramienta poderosa para el avance de la investigación sobre nutrición acuática. Sin embargo, pocos estudios han investigado exhaustivamente la respuesta de la microbiota del huésped a los oligoelementos. Este estudio examinó el papel de la suplementación con cobre orgánico en la promoción de la salud del camarón blanco de cultivo (Penaeus vannamei) desde la perspectiva de la microbiota.
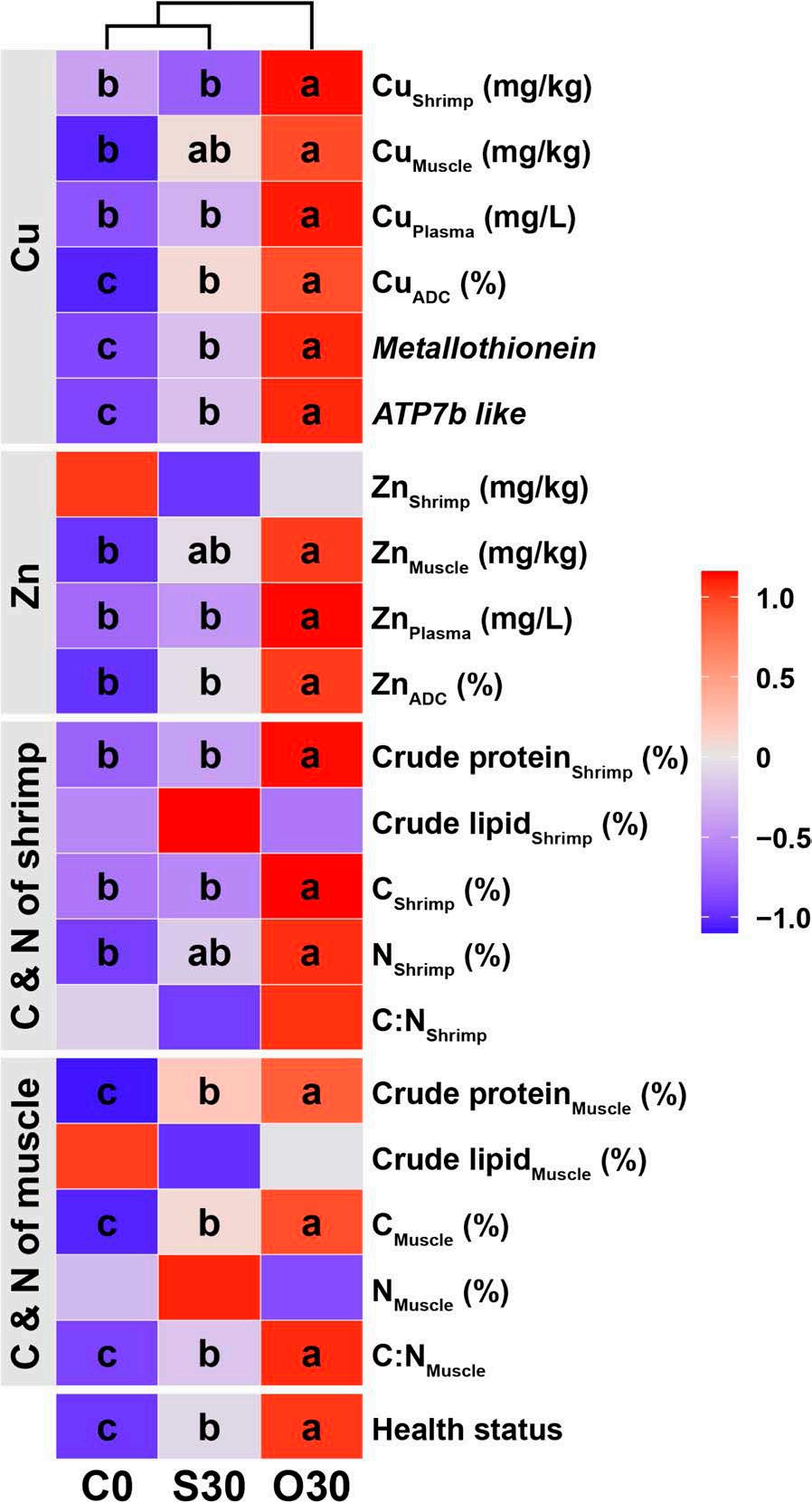
Resultados
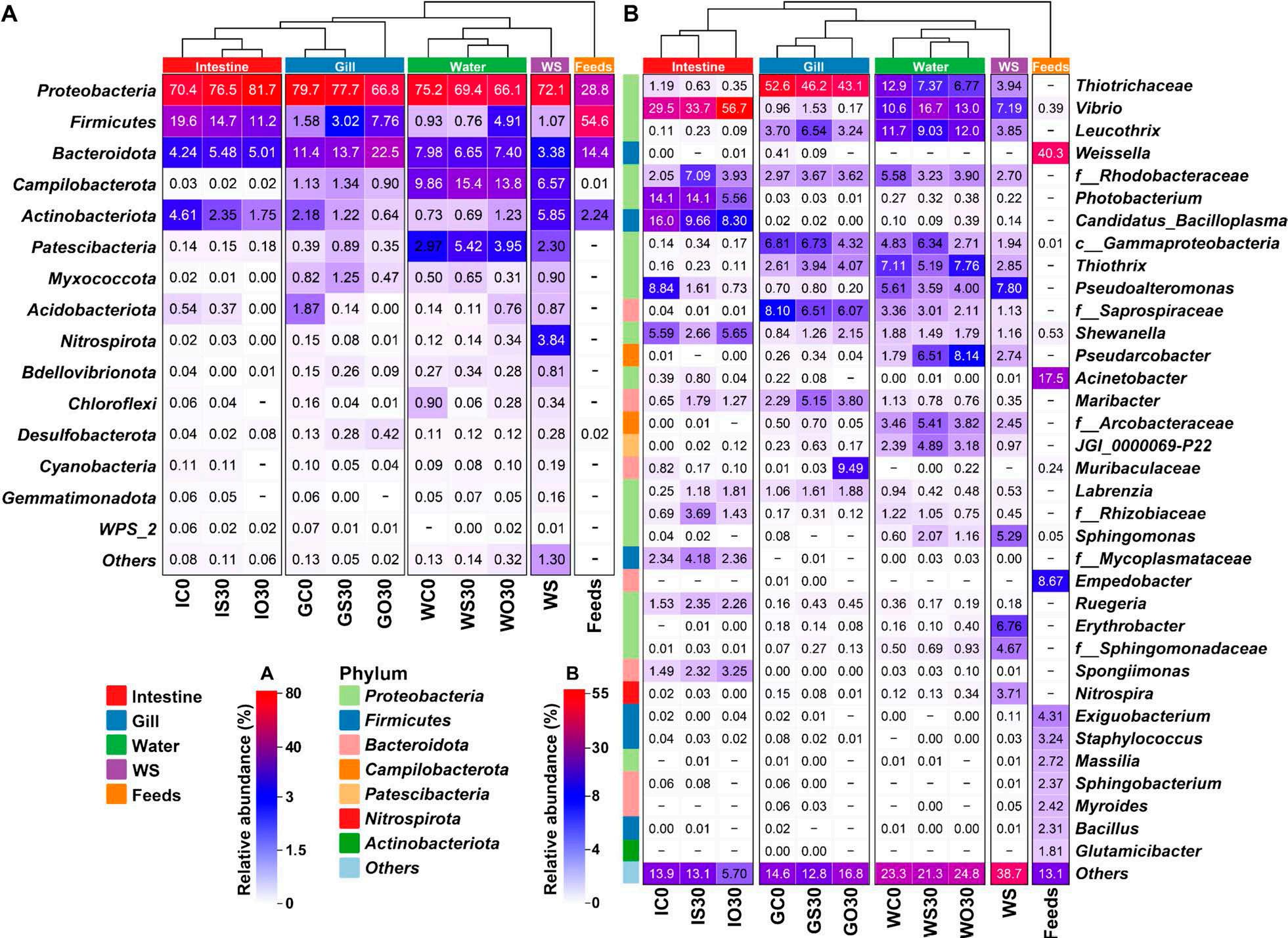
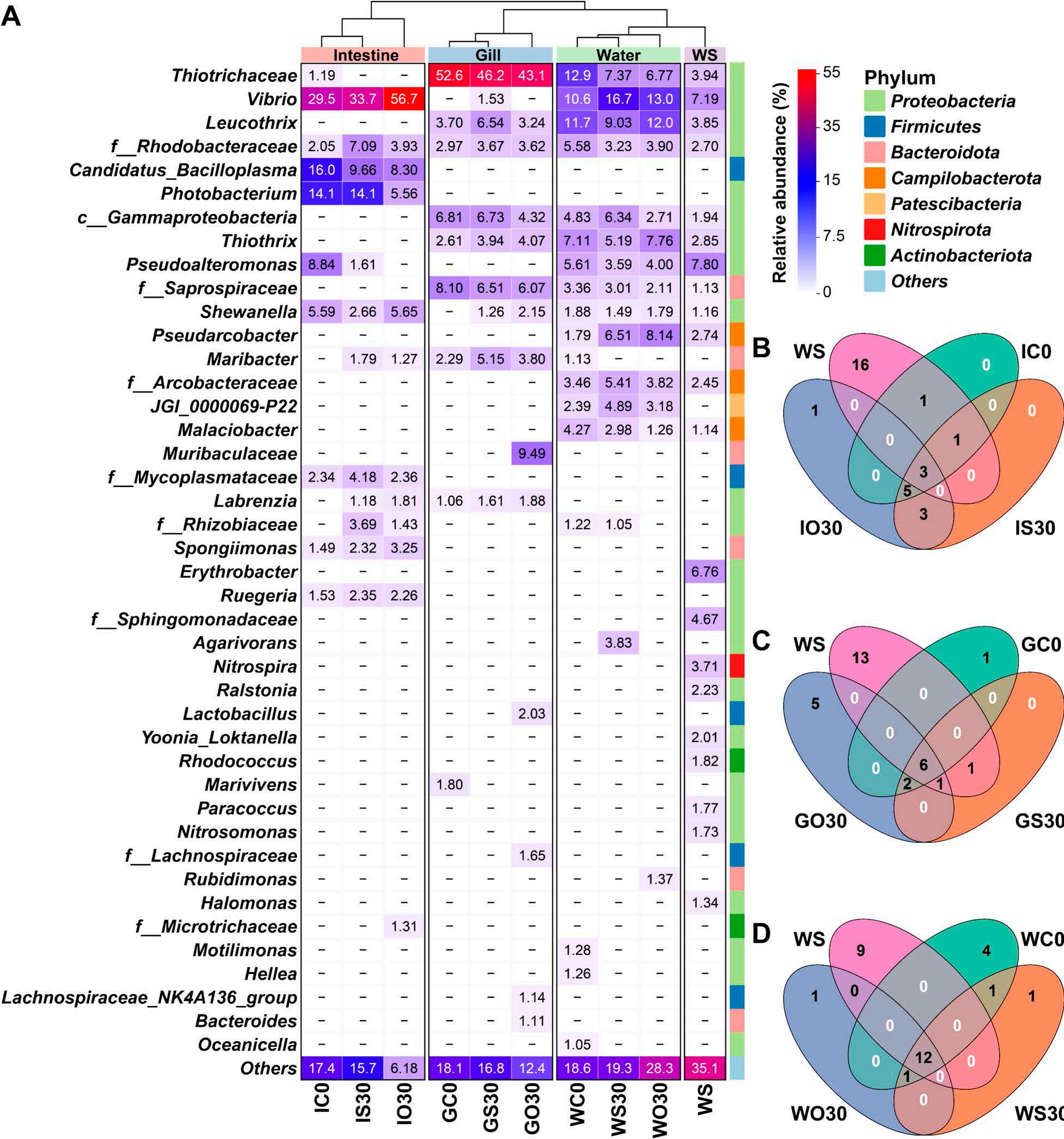
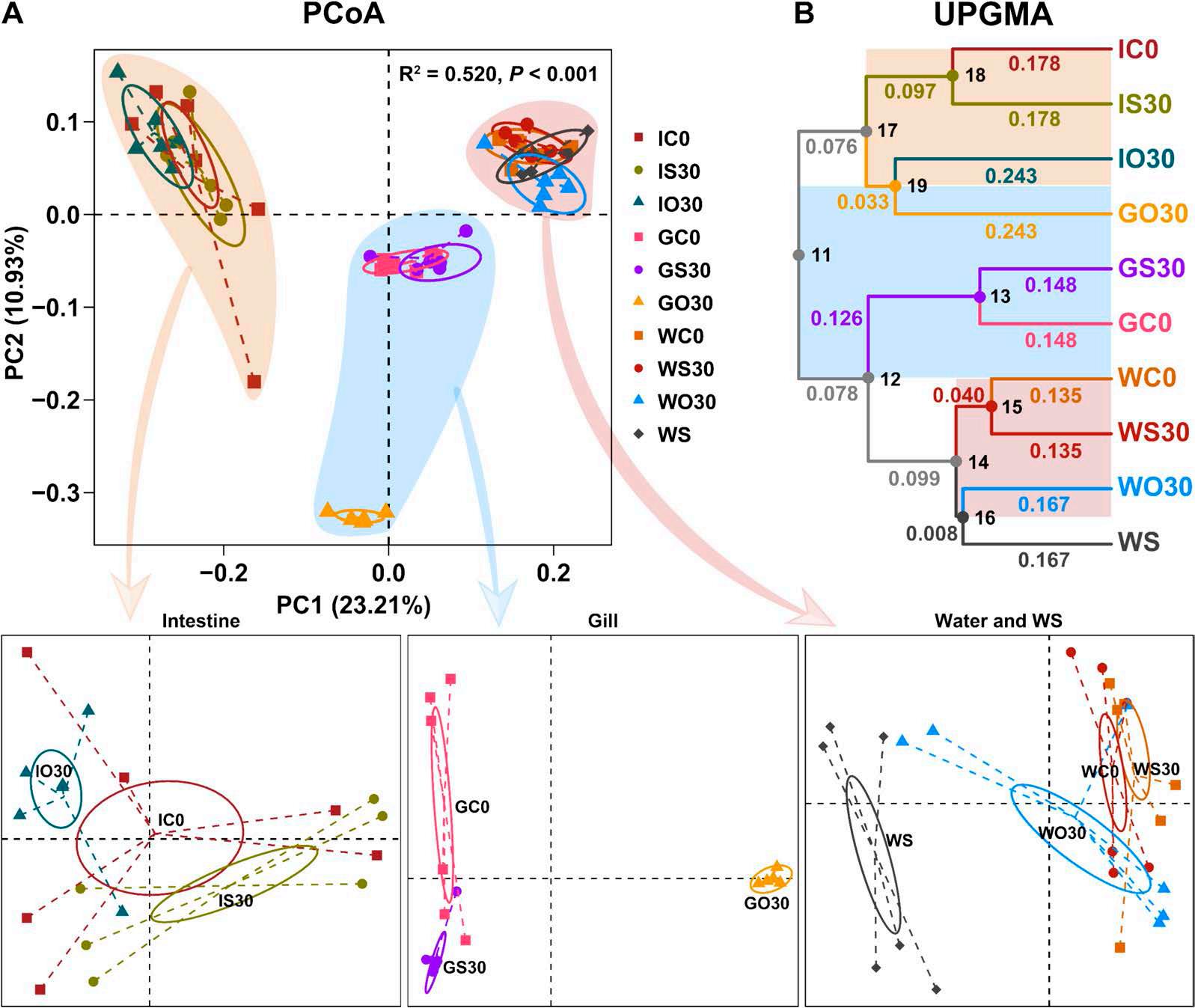
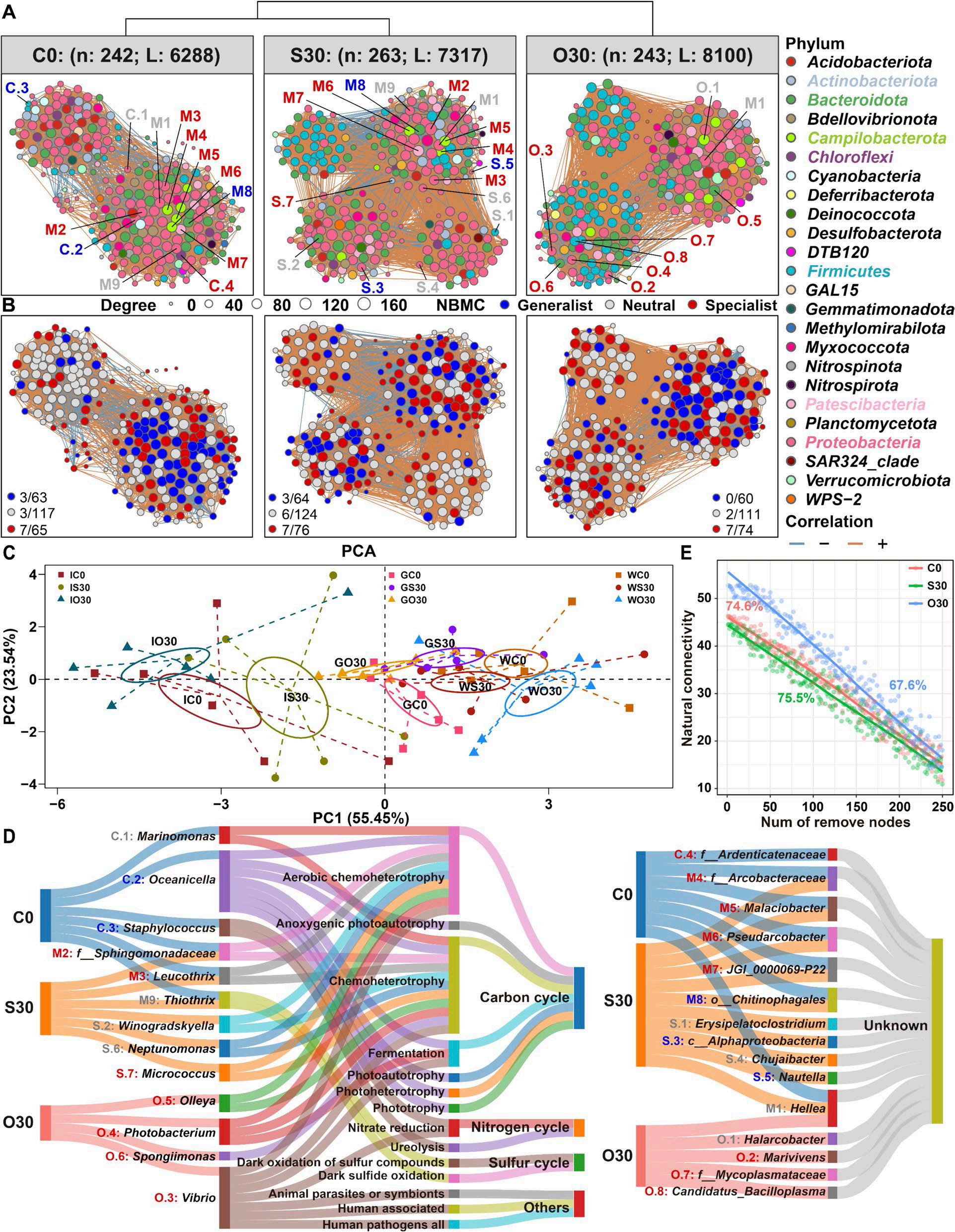
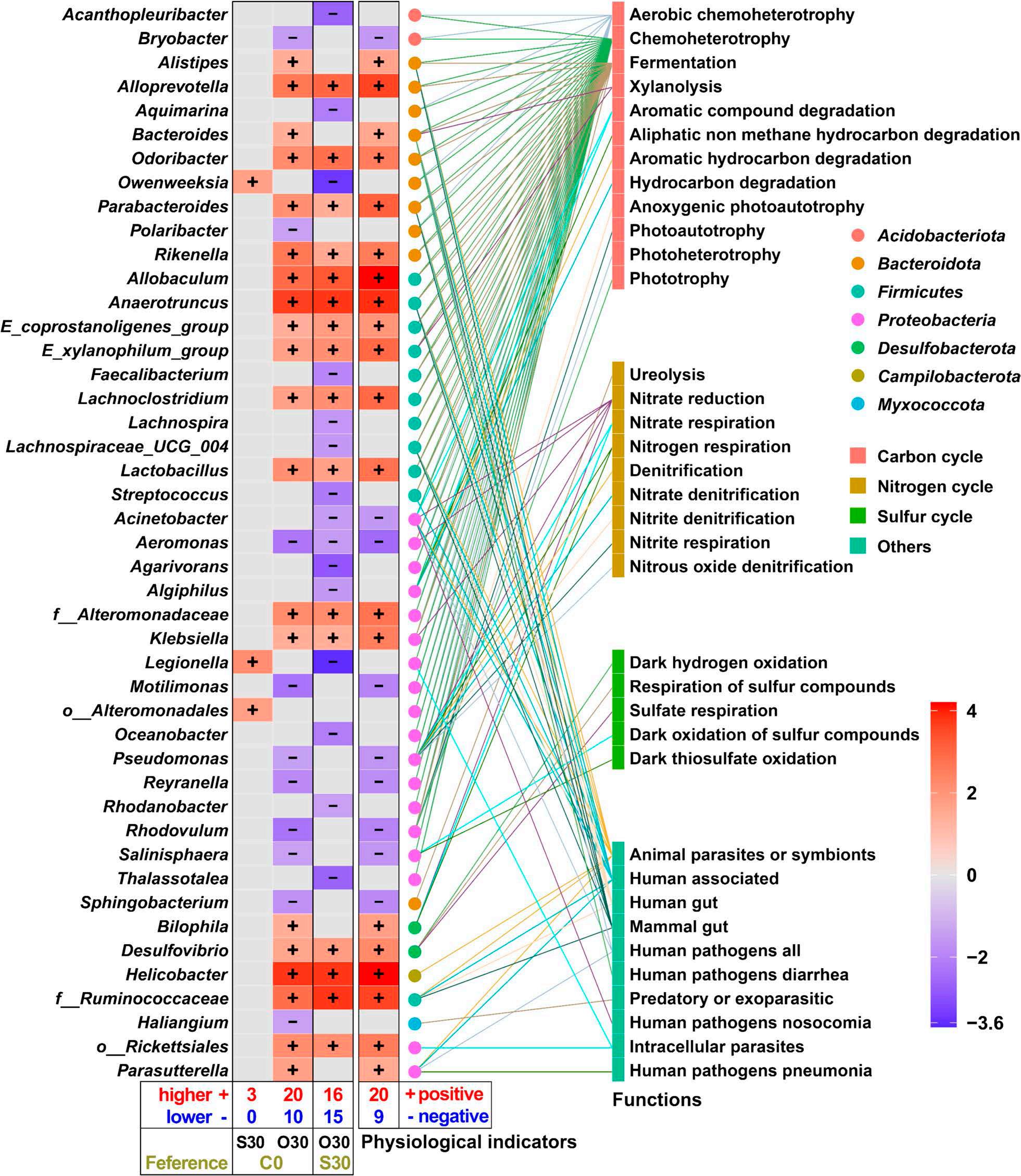
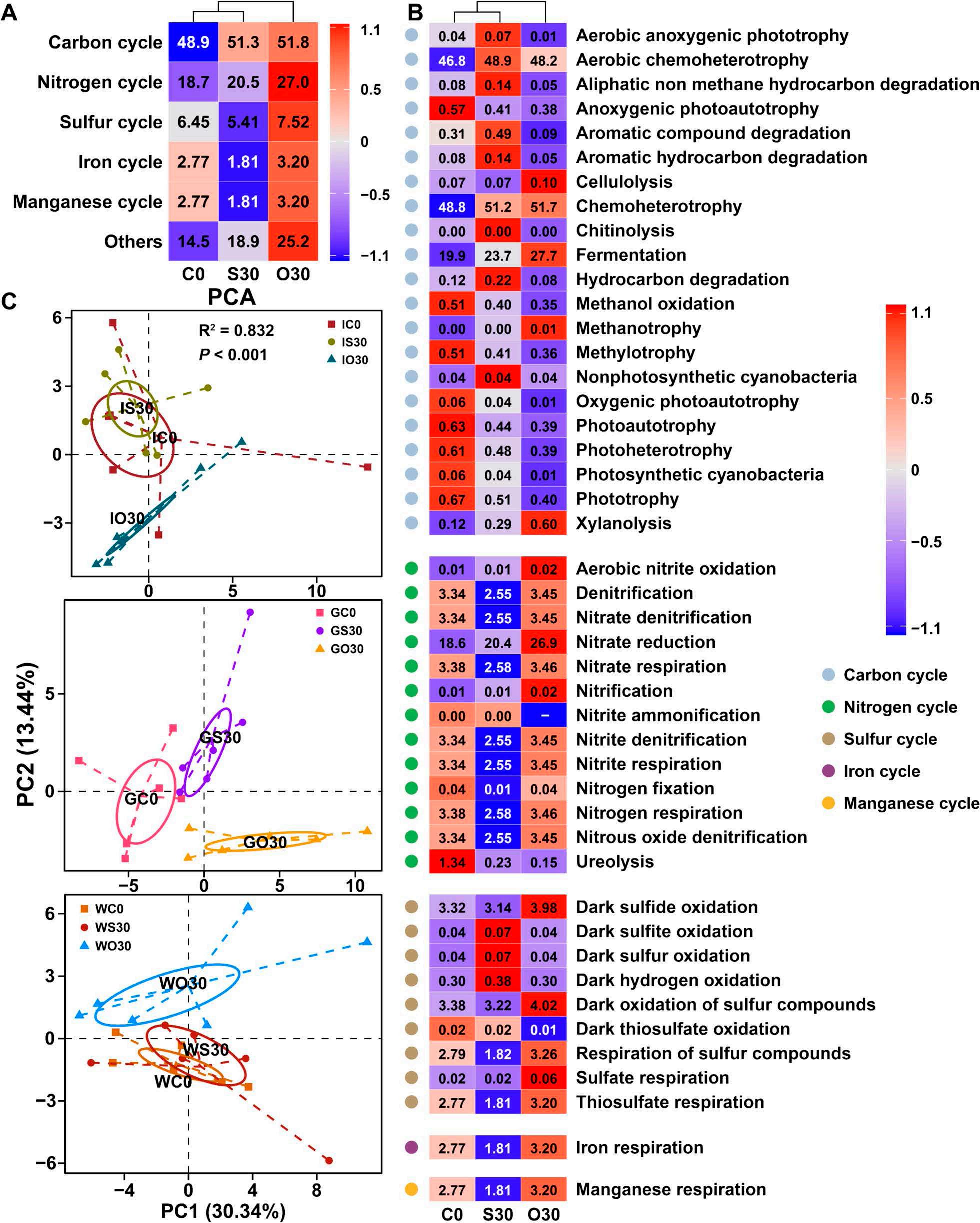
En un ensayo de alimentación de 8 semanas, los camarones fueron alimentados con dietas suplementadas sin cobre, 30 mg/kg de cobre inorgánico (CuSO4 · 5H2O) o cobre orgánico (Cu-proteinato). Se determinaron los coeficientes de digestibilidad aparente de cobre y zinc, junto con la asimilación de carbono y nitrógeno. La región V3-V4 del gen 16S rRNA fue secuenciada de alimentos, intestinos, branquias y muestras de agua. Los camarones que se alimentaron con la dieta de cobre orgánico demostraron un estado fisiológico más saludable, coeficientes de digestibilidad aparente más altos tanto de cobre como de zinc, así como una mayor acumulación de cobre, zinc, carbono y nitrógeno. El grupo de cobre orgánico exhibió una diversidad microbiana distintiva y una red de co-ocurrencia microbiana más compleja, caracterizada por una conectividad natural mejorada y robustez. Los taxones clave, incluidos Vibrio, Candidatus_Bacilloplasma y Photobacterium, contribuyeron a la estabilidad de la red. Los taxones asociados con el metabolismo de nutrientes, incluyendo Butyricicoccus, Lactobacillus y géneros de la familia Lachnospiraceae, Prevotellaceae, Rikenellaceae y Ruminococcaceae, se enriquecieron significativamente, correlacionándose bien con perfiles nutricionales mejorados. En concordancia, la anotación funcional reveló que el grupo de cobre orgánico exhibió mayores abundancias de módulos funcionales asociados con el metabolismo de nutrientes y energía como el ciclo del carbono y el nitrógeno. Además, la presión selectiva del huésped dio forma a la composición única de la microbiota en el intestino y las branquias, que difería del agua circundante y la fuente de agua, con la microbiota branquial sirviendo potencialmente como un puente de
transición que da forma a la intestinal.
Conclusiones
En este estudio, una microbiota hospedadora más estable, taxones metabolizadores de nutrientes enriquecidos y un ciclo ecológico mejorado ofrecen una posible estrategia para el desarrollo de alimentos acuícolas innovadores. Nuestros hallazgos ofrecen nuevas perspectivas, centradas en la microbiota, sobre el papel del cobre orgánico en el cultivo saludable del camarón.
Fondo
El camarón blanco (Penaeus vannamei) representa una fuente crucial de proteína de alta calidad y constituye un componente significativo de la producción mundial de mariscos, representando más del 50% de la producción mundial de crustáceos. En particular, los costos de alimentación representan más de la mitad de los gastos totales de producción [1]. La composición nutricional de los alimentos influye directamente en la salud del camarón, la calidad de la carne y el rendimiento de la producción. Entre los nutrientes esenciales, los minerales traza desempeñan papeles particularmente vitales en el crecimiento del camarón, la reproducción, la función inmune y el metabolismo energético, actuando como componentes o activadores de enzimas y hormonas para apoyar los procesos fisiológicos esenciales [2]. La creciente utilización de proteínas de origen vegetal en alimentos acuícolas ha aumentado la dependencia de minerales traza suplementados [3]. Si bien los minerales de sulfato inorgánico han sido tradicionalmente la principal opción de suplementación [4], los minerales traza orgánicos (OTM) son cada vez más reconocidos como una alternativa superior debido a su mayor biodisponibilidad y beneficios inmunológicos para el camarón. Esta ventaja es particularmente significativa en sistemas de cultivo de camarón intensivos y sostenibles, donde los OTM contribuyen a una mejor gestión de la calidad del agua [5]. La microbiota intestinal saludable apoya la absorción de nutrientes, la resistencia al estrés y la homeostasis energética en el camarón [6, 7, 8]. Como los factores nutricionales influyen significativamente tanto en el huésped como en su microbiota intestinal asociada [6], comprender estas interacciones es esencial para optimizar la
salud y la producción del camarón. Si bien la importancia de los minerales traza en la fisiología del camarón está bien establecida, la investigación que examina específicamente su impacto en la microbiota del camarón sigue siendo limitada. Nuestros estudios previos han demostrado que los orgánicos de zinc y manganeso promueven perfiles de microbiota intestinal más favorables en el camarón [9, 10]. El cobre, un mineral traza particularmente esencial para los crustáceos, es crucial para varias funciones fisiológicas [5]. Aunque nuestra investigación previa ha establecido los beneficios fisiológicos del cobre orgánico en el camarón blanco [11], sus efectos en la microbiota intestinal necesitan mayor exploración. Investigar estos efectos es esencial para desbloquear el potencial del cobre orgánico en la mejora de las interacciones huésped-microbiota y la mejora de la salud del camarón.
Además de las que existen en el intestino, las comunidades microbianas en las branquias y el agua circundante también están estrechamente asociadas con la salud del huésped [12, 13]. A diferencia de los animales terrestres expuestos principalmente al aire, los animales acuáticos están expuestos a cargas microbianas mucho más altas en su entorno acuático [12]. Los estudios en peces han demostrado que el entorno acuático puede influir en la composición de la microbiota intestinal [14, 15], al tiempo que revelan la especificidad de las comunidades y funciones microbianas en diferentes sitios del cuerpo (por ejemplo, branquias, intestino y piel) [12, 16]. En el camarón, las interacciones de la microbiota ocurren principalmente en el intestino, las branquias y el agua circundante. Un estudio en camarón blanco identificó la función inmunomoduladora de la microbiota branquial y las diferencias entre las comunidades de microbiota de agua de mar y branquial [17]. Sin embargo, a pesar de estos hallazgos, sigue faltando una investigación exhaustiva sobre las interacciones entre la microbiota intestinal, branquial y acuática del camarón y sus relaciones con la salud fisiológica del huésped.
El avance de las herramientas bioinformáticas ha permitido una interpretación más completa y profunda de los datos de secuenciación de amplicones. Sin embargo,
los investigadores se han encontrado con nuevos desafíos que podrían comprometer la precisión analítica. Uno de estos problemas se refiere a la existencia de la microbiota alimentaria. Aunque estos fragmentos de la microbiota alimentaria no significan bacterias vivas, son capaces de persistir en el intestino después de la ingestión y podrían aparecer en los resultados de la secuenciación. Estos fragmentos pueden ser identificados erróneamente como microbiota asociada al huésped, lo que podría sesgar el análisis. Este fenómeno se ha informado en el salmón del Atlántico (Salmo salar) [18, 19], donde el predominio de Lactobacillus en la digesta intestinal distal podría resultar de los fragmentos de ADN bacteriano que se originan en el alimento [19]. Por lo tanto, la realización de análisis de la microbiota alimentaria en experimentos de acuicultura puede proporcionar conclusiones más cautelosas y objetivas. Sin embargo, las descripciones de esta práctica son actualmente limitadas.
Considerando estas lagunas de conocimiento, en este estudio, pretendemos (1) investigar cómo el cobre orgánico regula la salud fisiológica del camarón y las interacciones de la microbiota en el intestino, las branquias y el agua circundante, (2) explorar los efectos ecológicos del cobre orgánico en la microbiota asociada al huésped, como el metabolismo del carbono y el nitrógeno, y (3) evaluar si la microbiota presente en el alimento para camarones blancos interfiere con el análisis de la microbiota del huésped. Nuestra hipótesis es que los efectos del cobre orgánico en la microbiota del huésped pueden ser más inesperados de lo que se pensaba anteriormente. Una evaluación exhaustiva de la microbiota asociada al huésped proporcionará nuevas perspectivas para futuras investigaciones en nutrición acuícola. Este estudio ofrece una perspectiva de la microbiota para optimizar las formulaciones de alimentos acuícolas y contribuye al desarrollo sostenible de la acuicultura.
Métodos
Diseños de experimentos y recolección de muestras
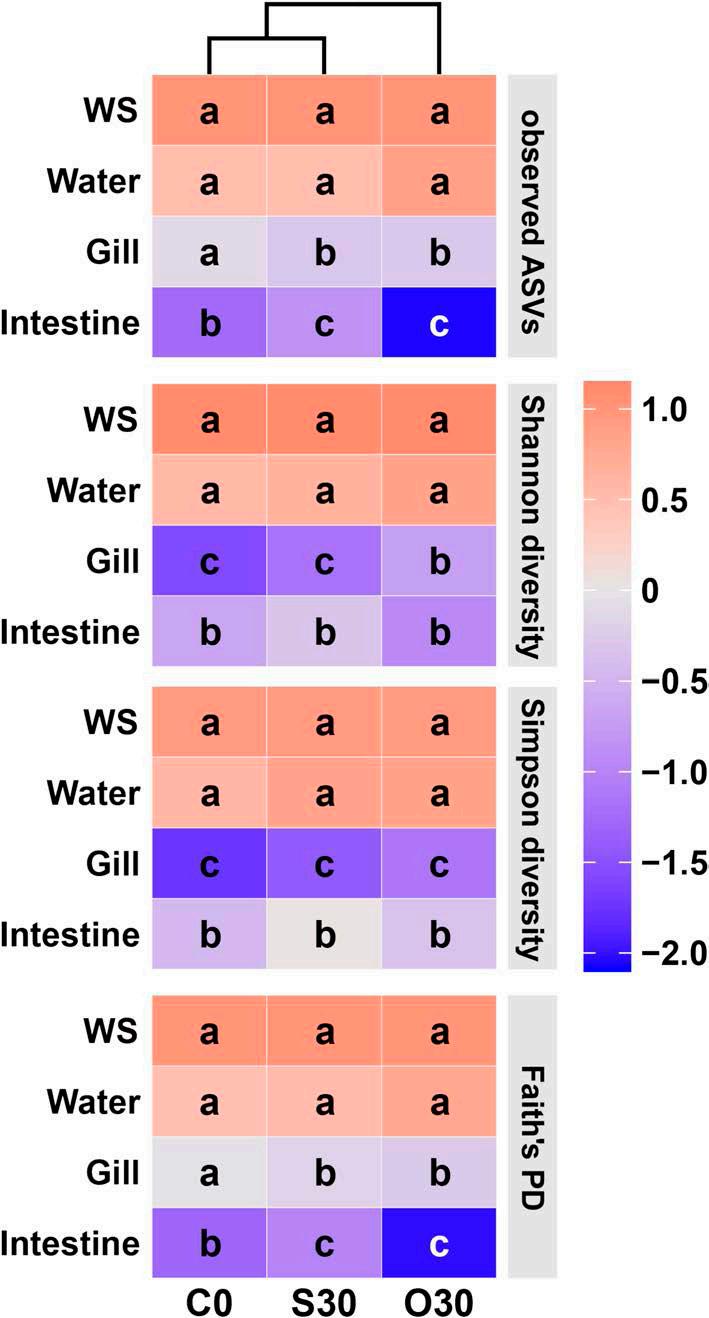
Las dietas experimentales y los ensayos de alimentación se describieron en Yang et al., 2023 [11]. Brevemente, se seleccionaron
muestras de tres dietas experimentales de ese estudio (Tabla S1): C0, sin suplementación de Cu; S30, 30 mg/kg de Cu inorgánico de CuSO 4 · 5H 2 O; y O30, 30 mg/ kg de Cu orgánico de Cu-proteinato. Cada dieta se asignó aleatoriamente a 3 tanques con 40 camarones/tanque (peso corporal inicial 5,29 g (DE = 0,03)). También citamos los resultados experimentales de estos tres tratamientos, incluyendo el contenido de proteína cruda y lípidos crudos de todo el camarón, los niveles de expresión genética de metalotioneína y ATP7b como en el hepatopáncreas, e indicadores relacionados con la salud (actividades enzimáticas antioxidantes e inmunológicas en plasma, expresiones genéticas antioxidantes e inmunológicas en el hepatopáncreas, tasa de supervivencia bajo estrés de 10 °C y recuentos de hemocitos).
La recolección de muestras siguió un protocolo con ciertas modificaciones [10, 18]. Brevemente, las heces se recolectaron diariamente después de una alimentación de 6 semanas. Se recolectaron muestras completas de camarón, músculo, branquias e intestino después de un intervalo de alimentación adicional de 2 semanas. Para cumplir con los requisitos de tamaño de muestra para la extracción de ADN, los intestinos y las branquias de cada 4 camarones se agruparon en tubos criogénicos estériles separados libres de ARNasa para la extracción, con 24 camarones utilizados por tratamiento para facilitar 6 análisis replicados. Si los intestinos contenían contenido, se retiraron con cuidado. Los intestinos y las branquias se congelaron en nitrógeno líquido y se almacenaron a -80 °C. Aproximadamente 50 g de las dietas de cada tratamiento se pulverizaron y se congelaron a -80 °C para la extracción de ADN. Para obtener muestras de microbiota del agua, se filtraron 1200 mL de agua de cada tanque a través de un filtro de polietersulfona hidrófilo Millipore Express® de 0,22 μm (GPWP04700; SigmaAldrich (Shanghai) Trading Co. Ltd., China) utilizando una bomba de vacío de diafragma. El filtro enriquecido con microbios se colocó en un tubo criogénico estéril libre de ARNasa de 10 mL con el lado superior hacia adentro. Luego, se congeló en nitrógeno líquido y se almacenó a - 80 °C. Se realizaron dos rondas de muestreo en 9 tanques. Las
Sample type Dietas Repeticiones × tratamiento
C0 S30 O30
Intestino IC0 IS30 IO30 6 × 3
Branquia GC0 GS30 GO30 6 × 3
Alimento FC0 FS30 FO30 1 × 3
Agua WC0 WS30 WO30 6 × 3 WSa WS 6 × 1
muestras de microbiota de la fuente de agua se recolectaron del pozo de alimentación de camarones, con recolecciones tomadas cada 30 minutos por un total de 6 veces. La muestra se recolectó alrededor de lámparas de alcohol encendidas, con instrumentos esterilizados por llama después de limpiarlos con alcohol al 75% antes de su uso. Los tipos de muestra y las etiquetas relevantes se resumieron en la Tabla 1, con un total de 63 muestras.
Análisis y visualización de indicadores fisiológicos
Los indicadores fisiológicos abarcan la acumulación de Cu y Zn en el camarón entero, músculo y plasma, así como el coeficiente de digestibilidad aparente (ADC) de Cu y Zn, la expresión de genes relacionados con el transporte de Cu, el contenido de proteína cruda, lípido crudo, C y N en el camarón entero y músculo, e indicadores relacionados con el estado de salud (por ejemplo, tasa de supervivencia bajo una prueba de estrés, enzimas y expresiones genéticas de procesos relacionados con la respuesta antioxidante e inmune). La determinación de Cu, Zn e itrio y el cálculo de ADC se llevaron a cabo siguiendo los procedimientos analíticos descritos por Yang et al. [10]. Para determinar las concentraciones de carbono (C) y nitrógeno (N), se empleó un analizador elemental (Elementar, Alemania) después de la liofilización de las muestras. Otros indicadores se evaluaron utilizando los métodos descritos por Yang et al. [11]. La mayoría de los indicadores fisiológicos se han descrito previamente en un artículo publicado [11]. En consecuencia, estos resultados se presentan como un mapa de calor en el artículo actual por razones de brevedad utilizando el paquete ggplot2 [20]. Las medias ± DE se proporcionan en la Tabla S2.
Extracción y secuenciación de ADN
El ADN genómico de muestras de intestino,
branquias y alimento se extrajo con el kit QIAamp PowerFecal® Pro DNA (51804, Qiagen, Alemania). Se establecieron dos controles negativos sin muestra (grupo NC-A) para el control de calidad durante el proceso de extracción de ADN. El ADN genómico de muestras de agua y de fuente hídrica se extrajo con el kit DNeasy® PowerWater® (14900–100-NF, Qiagen, Alemania). De igual forma, se establecieron dos controles negativos sin muestra (grupo NC-B). El control de calidad del ADN, la genoteca y la secuenciación (incluidos los controles negativos) fueron realizados por Novogene Genomics Technology Co., Ltd., Pekín, China, en la plataforma Illumina NovaSeq.
Procesamiento de datos de secuenciación La región V3–V4 del gen ARNr 16S se amplificó utilizando los cebadores 341F (5′–CCTAYGGGRBGCASCAG–3′) y 806R (5′–GGACTACNNGGGTATCTAA–3′). El procesamiento y análisis de los datos de secuencia se realizó utilizando el software QIIME 2 con algoritmos DADA2 [21]. El flujo de trabajo de análisis incluyó la demultiplexación de lecturas emparejadas sin procesar por sus códigos de barras únicos, el recorte de secuencias de cebadores directos (17 pb) e inversos (19 pb), la fusión de lecturas emparejadas y el filtrado de lecturas de baja calidad. Este proceso generó la tabla de variantes de secuencia de amplicón (ASV) y la tabla de secuencias características. Luego, se realizó la clasificación taxonómica utilizando la base de datos Silva 138 [22]. Después de obtener la tabla ASV y las asignaciones de taxonomía, los ASV identificados como Archaea, Unassigned, Chloroplast y Mitochondria, y los ASV con una frecuencia de menos de 3/63 (excluyendo los controles negativos) se filtraron. Los ASV que no se pudieron asignar al nivel de género se designaron con una letra de prefijo que indica el rango taxonómico más alto alcanzado, como “ c__ “ para clase, “ o__ “ para orden y “ f__ “ para familia.
Tabla 1. Tipo de muestra de secuenciación e información del tratamiento
Los ASV contaminantes también se filtraron utilizando el paquete decontam, basado en los principios: las contaminaciones tienden a ser inversamente proporcionales a la concentración de ADN bacteriano en las muestras y tienden a encontrarse en los controles negativos [23].
La cuantificación del ADN bacteriano en muestras de ADN genómico se realizó mediante qPCR, haciendo referencia a estudios previos con ciertas modificaciones [24, 25]. En resumen, el ensayo qPCR se realizó en el sistema de detección de PCR en tiempo real CFX96 Touch (Bio-Rad, Hercules, EE. UU.) en un volumen de 20 μL: 2 μL de muestra de ADN diluida (2,5 ng/μL), 10 μL de TaKaRa Taq™ HS Low DNA (2 ×, R090A, Takara Biomedical Technology (Beijing) Co., Ltd., China), 1 μL de SYBR Green I (20 ×, SL2190, Coolaber Science & Technology, Beijing, China), 0,6 μL de cebador directo (10 μM, 5′–CCATGAAGTCGGAATCGCTAG–3′), 0,6 μL de cebador inverso (10 μM, 5′–GCTTGACGGGCGGTGT–3′), 5,8 μL de dH 2 O para ADN bajo. Se incluyeron estándares de ADN bacteriano y un control sin plantilla (E2006–2, Zymo Research Corp, CA, EE. UU.) en cada reacción de placa de qPCR. El programa de qPCR fue el siguiente: 35 ciclos de 5 s a 94 °C y 20 s a 65 °C. Para garantizar las especificaciones del producto, se utilizó la curva de fusión después de la amplificación. El valor de Cq del control sin plantilla se obtuvo después de 35 ciclos, lo que refleja la ausencia de contaminación de ADN durante el proceso de qPCR. Las concentraciones del gen ARNr 16S se muestran en la Tabla S3.
Visualización de datos de secuenciación
Después del filtrado, el árbol filogenético de ASV limpios se construyó en Wekemo Bioincloud [26]. Para evaluar la profundidad de la secuenciación, se trazaron diagramas de caja de acumulación de especies, abundancia de rango y curvas de rarefacción utilizando los paquetes vegan, doBy, BiodiversityR y ggplot2 [27, 28, 29]. Los paquetes ggplot2 y ggalluvial se utilizaron para visualizar las 10 características principales, incluidos el filo y el género [30]. Las características se trazaron por separado en función de los tipos de muestra, que incluyeron intestino, branquias, agua, fuente de agua y alimentos, y luego se agregaron en un mapa de calor utilizando el paquete
ggplot2 . Las características principales (nivel de género) se identificaron como aquellas con una prevalencia de más del 80% y abundancias relativas promedio (Ave) de más del 1% en cada tratamiento y se visualizaron como un mapa de calor utilizando el paquete ggplot2 y diagramas de Venn utilizando la plataforma CNSknowall (https://cnsknowall. com/#/HomePage).
Los índices de diversidad alfa de ASV observados, diversidad de Shannon, diversidad de Simpson y diversidad filogenética de Faith (PD de Faith) se calcularon utilizando los paquetes picante y vegan [31]. Estos índices se agregaron luego en un mapa de calor utilizando el paquete ggplot2. Las medias ± DE y las fórmulas de cálculo de la diversidad de Shannon y la diversidad de Simpson se proporcionan en la Tabla S4. La matriz de distancia de diversidad beta UniFrac no ponderada se calculó utilizando los paquetes ape y phyloseq y se visualizó como gráficos de análisis de coordenadas principales (PCoA) y agrupamiento mediante el método de grupo de pares no ponderado con media aritmética (UPGMA) utilizando los paquetes ggplot2, ggtree y treeio [32, 33, 34, 35].
La red de co-ocurrencia a nivel de género fue visualizada usando los paquetes ggplot2 y ggClusterNet de acuerdo con las instrucciones proporcionadas [36]. Antes del análisis de la red de co-ocurrencia, las características (nivel de género) con menos del 50% de prevalencia y 0.01% Ave en cada tratamiento fueron excluidas. Solo el coeficiente de correlación | r |> 0.6 y P < 0.05 fueron mostrados al construir la red de co-ocurrencia. La amplitud de nicho de la comunidad microbiana (NBMC) fue calculada haciendo referencia a Yan et al. [37]. La función ZiPiPlot y la función Hub_score fueron utilizadas para identificar características clave (también llamadas taxones keystone) en la red. El diagrama de Sankey de características clave fue visualizado usando la plataforma ChiPlot (https://www.chiplot.online). Las funciones Robustness.Random.removal y natural.con. microp fueron utilizadas para evaluar la capacidad de resistir perturbaciones de la red de co-ocurrencia. Las funciones potenciales se predijeron utilizando la base de datos de anotación
funcional de taxones procariotas (FAPROTAX) (v 1.2.7) en Python (v 3.10.12) corriendo bajo Linux (para anotar las funciones ecológicas correspondientes a las características) [38] y utilizando el paquete Tax4Fun2 en R (para obtener la abundancia de la vía potencial) [39].
Análisis estadístico
Como se ilustra en la Fig. S1, los indicadores relacionados con el estado de salud mostraron una alta correlación. En consecuencia, se realizó un análisis de componentes principales (PCA) utilizando el paquete factoextra en R (v 4.2.3) [40, 41], y se empleó el PC1 para la prueba de asociación, dado que estos indicadores se correlacionaron negativamente con el PC1 del PCA. Los métodos de análisis estadístico empleados en la Tabla S2 (sección de indicadores fisiológicos) se alinearon con los descritos previamente en el artículo [11]. Brevemente, se utilizó un análisis de varianza unidireccional para evaluar la significancia estadística de las diferencias utilizando el paquete agricolae [42]. Las estadísticas se consideraron significativas cuando P < 0,05. El modelo lineal de efectos mixtos con el paquete lme4 se utilizó para comparar los índices de diversidad alfa [43]. El efecto fijo fue la información del grupo, como se muestra en la Tabla 1. El efecto aleatorio fue la distribución de los tanques. Se utilizaron ANOVA de tipo III y aproximaciones de Kenward-Roger para probar los efectos fijos con el paquete lmerTest [44]. Se realizaron comparaciones múltiples si el efecto fijo era significativo utilizando el paquete emmeans [45]. P < 0,05 se consideró significativo. Se realizó una prueba de análisis de varianza multivariado permutacional (PERMANOVA) utilizando la función adonis2 dentro del paquete vegan
Las características asociadas significativas (nivel de género) y la detección de vías predictivas diferenciales (resultados de Tax4Fun2) se llevaron a cabo utilizando el paquete maaslin2 [46]. La presencia de una diferencia se consideró estadísticamente significativa cuando Q < 0,25 (P –ajuste). Al realizar un análisis de características diferenciales, las características con menos del 50% de prevalencia y 0,01% de Ave en cada tratamiento no se someterían a análisis. Las dietas (C0, S30 y O30) y los
indicadores fisiológicos fueron el efecto fijo. Como se ilustra en la Fig. S2, los indicadores fisiológicos exhibieron una alta correlación excepto por el contenido de Zn y lípidos crudos y la relación C a N del camarón entero, así como el contenido de lípidos crudos y N del músculo, que no demostraron cambios significativos entre todos los grupos. Se empleó el − PC1 para la prueba de asociación, dado que estos indicadores se correlacionaron negativamente con el PC1 del PCA. Al realizar un análisis de la vía diferencial, el efecto fijo fue la información del grupo (IC0, IS30 y IO30; GC0, GS30 y GO30; WC0, WS30 y WO30), como se muestra en la Tabla 1, excepto para los feeds y las muestras WS.
Resultados
Conclusiones de estudios previos Los resultados de nuestras investigaciones anteriores indicaron que, en comparación con el cobre inorgánico, los camarones alimentados con una dieta de cobre orgánico exhibieron mayores parámetros antioxidantes e inmunes, recuentos más altos de hemocitos y una mayor tasa de supervivencia bajo estrés de 10 °C [11].
Indicadores fisiológicos
Los camarones del grupo S30 mostraron indicadores fisiológicos similares a los del grupo control (C0) (Fig. 1). Si bien el grupo S30 mostró algunas mejoras fisiológicas, el grupo O30 demostró un rendimiento superior en múltiples aspectos: mayor ADC de Cu y Zn, mayor acumulación de Cu y Zn, y mayor eficiencia en el transporte de Cu. Además, el grupo O30 logró una mayor retención de C y N y mostró un mejor estado de salud general.
Control de calidad
El análisis de contaminación reveló 165 variantes de secuencia de amplicón (ASV) en muestras intestinales, branquiales y de alimento, lo que representa 67.703 lecturas. Estas secuencias contaminantes se anotaron predominantemente como Sphingomonas, Methylobacterium ethylorubrum, Vibrio, JGI_0000069-P22, c__ Gammaproteobacteria, entre otras (Tabla S 5). Las muestras de agua y fuente de agua contenían una mayor cantidad de ASV contaminadas (412), lo que representa 113.120 lecturas, con taxones predominantes queincluían Alcanivorax, Escherichiaella, Te-
nacibaculum, Enterococcus, Faecalibacterium, Lacihabitans y otros (Tabla S6).
Después de la eliminación del contaminante, el conjunto de datos final comprendió 2.449 ASV con 2.925.425 lecturas, que representan 33 filos, 68 clases, 194 órdenes, 303 familias, 554 géneros y 235 especies. Las muestras de control negativo mostraron un predominio de Proteobacteria, Bacteroidota, Campilobacterota y Firmicutes a nivel de filo, mientras que Thiotrichaceae, Ralstonia, c__Gammaproteobacteria, Leucothrix, f__Saprospiraceae, f__Rhodobacteraceae, Vibrio, Thiothrix, Sphingomonas y Maribacter fueron los géneros dominantes (Fig. S 3). Además, el diagrama de caja de acumulación de especies, el diagrama de abundancia de rangos y las curvas de rarefacción confirmaron que la tabla ASV limpia final proporcionó un volumen de datos suficiente para el análisis posterior (Fig. S4).
Taxonomía y características dominantes Las 10 características principales (filo y género) mostraron variación en su composición entre los diferentes tipos de muestra; sin embargo, dentro de cada tipo de muestra, la composición de estas características fue similar entre los subgrupos, como se ilustra en la Fig. S 5. En consecuencia, las características dominantes se agregaron en mapas de calor que abarcan los niveles de filo (Fig. 2 A) y género (Fig. 2 B) para cada tratamiento.
Los alimentos exhibieron una composición de filo distinta, que comprendía solo 6 filos (Firmicutes, Proteobacteria, Bacteroidota, Actinobacteriota, Desulfobacterota y Campilobacterota). El género dominante (por ejemplo, Weissella, Acinetobacter, Empedobacter) difirió notablemente del observado en los otros tipos de muestra. La agrupación de filos y géneros también destacó la divergencia sustancial de los alimentos de los grupos restantes (Fig. 2). Además, el 25% de los géneros más abundantes en los alimentos estaban todos presentes en baja abundancia en el camarón, excepto Vibrio y Shewanella, que estaban presentes en mayor abundancia en el camarón (Fig. S6).
A nivel de filo, Proteobacteria fue el más abundante en todos los tipos de muestra
Fig 1. Efectos del cobre orgánico en la composición nutricional y la salud fisiológica del camarón blanco. Se aplicó la normalización de la puntuación Z a cada índice al construir los mapas de calor. La agrupación en el mapa de calor se realizó mediante la distancia de Bray-Curtis con el método de ligamiento promedio. Las medias ± DE se proporcionan en la Tabla S 2. a, b, c Los valores que no comparten la misma letra en superíndice de cada fila son significativamente diferentes (P < 0,05). ADC, coeficiente de digestibilidad aparente; similar a ATP7b, similar a la ATPasa 2 transportadora de cobre; C, carbono; Cu, cobre; N, nitrógeno; Zn, zinc. El texto en cursiva indica los resultados de qPCR. Los indicadores relacionados con el estado de salud se ilustraron en la Fig. S 1. Información del grupo: C0, sin suplementación de Cu; S30, 30 mg/kg de suplementación de Cu inorgánico; O30, 30 mg/kg de suplementación de Cu orgánico.
(intestino, branquias, agua y WS). El segundo filo más prevalente varió según el tipo de muestra: Firmicutes en el intestino, Bacteroidota en las branquias, Campilobacterota en el agua, tanto Campilobacterota como Actinobacteriota en el WS (Fig. 2 A). El género dominante también
exhibió variación entre los tipos de muestra (Fig. 2 B). En el intestino, Candidatus_Bacilloplasma y Photobacterium fueron los géneros más prevalentes. En las branquias, los géneros dominantes incluyeron f__Saprospiraceae, Maribacter y Muribaculacea f__Arcobacteraceae y JGI_0000069-P22 fueron dominantes en el agua, mientras que Erythrobacter, Sphingomonas, f__Sphingomonadaceae y Nitrospira dominaron el WS. Algunos géneros mostraron una amplia distribución en múltiples tipos de muestra. Vibrio y Pseudoalteromonas fueron abundantes en el intestino, el agua y el agua. f__Rhodobacteraceae fue predominante en el intestino, las branquias y el agua. Thiotrichaceae, Leucothrix y Thiothrix fueron abundantes en las branquias, el agua y el agua. c__Gammaproteobacteria fue dominante en las branquias y el agua. Shewanella fue dominante en el intestino y las branquias.
Características principales
Se obtuvo un total de 42 características principales entre todos los grupos, que incluyeron casi todos los géneros dominantes (Fig. 3). Con base en las disimilitudes de Bray-Curtis, el análisis de características principales reveló una agrupación relacionada con el tejido y la dieta, con el agua y el WS agrupándose inicialmente juntos, seguidos de agruparse con las branquias y finalmente con el intestino. Además, dentro de los tipos de muestra de intestino, branquias y agua, los grupos C0 y S30 muestran una composición similar de características principales, que posteriormente se agruparon con el grupo O30.
Además, el intestino y las branquias contenían las características centrales con una abundancia relativa abrumadoramente dominante, con el Vibrio en el intestino excediendo el 29,5% (Ave) y las Thiotrichaceae en las branquias sobrepasando el 43,1% (Ave). Por el contrario, el WS se caracterizó por diversas características centrales que se distribuyeron uniformemente, con Ave que oscilaba entre 1,13 y 7,80%. La agregación de algunas características centrales también estaba presente en el agua, incluyendo Thiotrichaceae, Vibrio y Leucothrix, con Ave que oscilaba entre 6,77 y 16,7% (Fig. 3 A).
Los diagramas de Venn indicaron que 3 características principales (Vibrio, f__Rhodo-
Fig 2 . Gráfico combinado de los 10 filos principales (A) y géneros de todos los grupos (B) en la Fig. S5. Los taxones no asignados a nivel de género están precedidos por una letra (c__, clase y f__, familia) que indica el nivel al que están asignados. La agrupación se realizó mediante la distancia de Bray-Curtis con el método de ligamiento promedio. Los números en las celdas representan la abundancia relativa promedio (%). ‘ − ‘ indica que la característica no existía en ese grupo. WS: fuente de agua. Los grupos de muestra se designaron utilizando prefijos para indicar los tipos de muestra: ‘I’ para muestras intestinales, ‘G’ para muestras branquiales y ‘W’ para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
bacteraceae y Shewanella) eran comunes al WS (3/21) y al intestino (3/14) (Fig. 3 B), 6 características principales (por ejemplo, Thiotrichaceae, Leucothrix y f__Rhodobacteraceae) eran comunes al WS (6/21) y a las branquias (6/16) (Fig. 3 C), y 12 características principales (por ejemplo, f__Saprospiraceae, Shewanella y Malaciobacter) eran comunes al WS (12/21) y al agua (12/20) (Fig. 3 D).
Diversidad alfa
Como se ilustra en la Fig. 4, dentro de los tipos de muestra de intestino, branquias y agua, no se encontraron diferencias significativas en los índices de diversidad alfa (ASV observados, diversidad de Shannon, diversidad de Simpson y PD de Faith) entre los tratamientos. Sin embargo, las diferencias entre los tipos de muestra revelaron la complejidad del entorno acuático. El agua y el WS mostraron índices de diversidad alfa similares (P > 0,05). Además, los ASV observados y el PD de Faith del intestino
exhibieron valores significativamente más bajos que las branquias, el agua y el WS (P < 0,05). Los valores medios observados de ASV fueron 203 (intestino) < 337 (branquias) < 530 (agua) < 598 (WS). Los valores medios de PD de Faith fueron 18,0 (intestino) < 28,9 (branquias) < 40,7 (agua) < 45,9 (WS). Las diversidades de Shannon y Simpson del agua y el WS fueron considerablemente más altas que las del intestino y las branquias. La diversidad de Shannon en las branquias de los grupos C0 y S30 fue significativamente menor que en el intestino. La diversidad de Simpson en las branquias fue sustancialmente menor que en el intestino (P < 0,05). Los valores medios de diversidad de Shannon fueron 23,9 (branquias) < 29,1 (intestino) < 100,0 (agua) < 131,3 (agua). Los valores medios de diversidad de Simpson fueron 5,5 (branquias) < 12,8 (intestino) < 35,7 (agua) < 40,0 (agua).
Diversidad beta
El gráfico de PCoA UniFrac no ponderado
SALUD
mostró una clara distinción entre los tipos de muestra (intestino, branquias, agua y WS) (R² = 0,520, P < 0,001). El análisis identificó tres grupos principales: (1) intestino (IC0, IS30 e IO30), (2) branquias (GC0, GS30 y GO30), y (3) agua y WS (WC0, WS30, WO30 y WS). Los subgráficos de PCoA ampliados demostraron que las comunidades de microbiota de los grupos C0, S30, O30 y WS formaron grupos distintos (Fig. 5 A). El análisis UPGMA indicó que las comunidades de microbiota de los grupos C0 y S30 mostraron mayor similitud en el intestino, las branquias y el agua (Fig. 5 B). Las pruebas PERMANOVA no revelaron diferencias significativas entre las comunidades de microbiota de los grupos IC0 e IS30 (P = 0,451), ni entre los grupos WC0 y WS30 (P = 0,129). Sin embargo, las comunidades de microbiota de las muestras de intestino, branquias y agua difirieron significativamente de las muestras de WS (P < 0,05). Además, se observaron diferencias significativas entre varios pares de grupos: IC0 frente a IO30, IS30 frente a IO30, GC0 frente a GS30, GC0 frente a GO30, GS30 frente a GO30 y WS30 frente a WO30 (P < 0,05) (Tabla S 7).
Redes de co-ocurrencia
Se seleccionaron un total de 390 características (pertenecientes a 24 filos, con una presencia predominante de Acidobacteriota, Bacteroidota, Firmicutes y Proteobacteria) para establecer redes de coocurrencia, que se emplearon para comparar las diferencias en las interacciones microbianas asociadas al huésped (intestino, branquias y agua circundante del camarón blanco) dentro de cada grupo dietético (C0, S30 y O30) y para identificar las características clave en la red de cada tratamiento. La varianza entre la red ER y la red subraya la validez de las redes específicas del grupo (Fig. S 7 A). Aunque la mayoría de las características dentro de cada tratamiento se correlacionaron positivamente, la distribución de los filos y NBMC de estas características en cada tratamiento reveló propiedades de red distintas (Fig. 6 A–C). El análisis de agrupamiento de los parámetros de la red reveló una estructura de red de coocurrencia única del grupo O30 (Fig. 6 A, B, Tabla 2). Los parámetros de red aumentaron en el orden de los grupos C0, S30 y O30, incluyendo enlaces totales (6288 < 7317 < 8100), enlaces positivos (5711 < 5882 < 7304), grado promedio (51,97 < 55,64 <
Fig 3. Mapa de calor (A) y diagramas de Venn (B - D) de las características principales del intestino, las branquias, el agua y el agua dulce (WS). Los taxones no asignados a nivel de género están precedidos por una letra (c__, clase y f__, familia) que indica su nivel asignado. La agrupación se realizó mediante la distancia de Bray-Curtis con el método de ligamiento promedio. Los números en las celdas representan la abundancia relativa promedio (%). ‘ − ‘ indica que la característica no fue clave en ese grupo. WS: fuente de agua. Los grupos de muestra se designaron mediante prefijos para indicar los tipos de muestra: ‘I’ para muestras intestinales, ‘G’ para muestras de branquias y ‘W’ para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
66,67), cercanía de centralización (0,250 < 0,268 < 0,290) y modularidad relativa (5,40 < 5,53 < 5,72). En contraste directo, la longitud de ruta promedio (1,51 > 1,35 > 1,27), el diámetro (3,90 > 3,17 > 3,10) y la intermediación de centralización (0,0668 > 0,0562 > 0,0510) de las redes disminuyeron progresivamente a través de los grupos C0, S30 y O30 (Tabla 2).
Utilizando los análisis ZiPiPlot (Fig. S7B) y hub_score (Tabla S8), se identificaron 28 características clave (taxones clave) y se
asignaron a los siguientes 7 filos: Actinobacteriota, Bacteroidota, Campilobacterota, Chloroflexi, Firmicutes, Patescibacteria y Proteobacteria. Entre estos, los grupos C0, S30 y O30 identificaron 13 características clave (3 generalistas, 3 neutrales y 7 especialistas), 16 (3 generalistas, 6 neutrales y 7 especialistas) y 9 (2 neutrales y 7 especialistas), respectivamente. Dentro de estos, el grupo C0 contenía 4 características clave únicas (f__Ardenticatenaceae, Marinomonas, Oceanicella y Staphylococcus), el grupo S30 contenía 7 características clave únicas (c__
Alphaproteobacteria, Chujaibacter, Erysipelatoclostridium, Micrococcus, Nautella, Neptunomonas y Winogradskyella), y el grupo O30 contenía 8 características clave únicas (Candidatus_Bacilloplasma, f__Mycoplasmataceae, Halarcobacter, Marivivens, Olleya, Photobacterium, Spongiimonas y Vibrio). Además, Hellea estaba presente en todos los grupos, mientras que 8 características clave adicionales (f__Arcobacteraceae, f__Sphingomonadaceae, JGI_0000069-P22, Leucothrix, Malaciobacter, o__Chitinophagales, Pseudarcobacter y Thiothrix) se compartían entre los grupos C0 y S30 (Fig. 6 A, B). El análisis FAPROTAX indicó que estas características clave abarcaban múltiples funciones ecológicas, como la quimioheterotrofia aeróbica, la quimioheterotrofia y la fermentación (ciclo del carbono), la reducción de nitratos (ciclo del nitrógeno) y la oxidación oscura de sulfuros (ciclo del azufre) (Fig. 6 D).
La conectividad natural de las redes en cada tratamiento disminuyó progresivamente con la eliminación de nodos. Sin embargo, el grupo O30 mantuvo una mayor conectividad natural en comparación con los grupos C0 y S30. Además, la reducción porcentual en la conectividad natural fue menor en el grupo O30 (67,6%) que en los grupos C0 (74,6%) y S30 (75,5%) (Fig. 6 E). El análisis de robustez de las redes en cada tratamiento muestra que, tras la eliminación aleatoria del 50% de los nodos, la robustez del grupo O30 (44,7%) sigue siendo mayor que la del grupo C0 (43,7%) y S30 (43,1%) (Fig. S7C).
Características diferenciales y sus funciones relacionadas
En general, 105 características diferenciales (nivel de género) se asociaron significativamente con las dietas y los indicadores fisiológicos del camarón blanco. De estas, 36 funciones ecológicas se anotaron en las 45 características diferenciales en la base de datos FAPROTAX (Fig. 7). Las funciones de las 60 características diferenciales restantes no se anotaron (Fig. S8). Específicamente, el análisis comparativo entre los grupos C0 y S30 reveló solo 7 características diferenciales. El grupo C0 mostró una abundancia relativa significativamente menor de Legionella, o__Alteromonadales y Owenweeksia en comparación con el grupo S30 (Fig. 7), mientras que mostró una abundancia rela-
tiva mayor de Chitinophaga, GAL15, Nautella y RB41 en comparación con los grupos S30 y O30 (Fig. S8). Por el contrario, cuando el grupo O30 se comparó con los grupos C0 y S30, se detectaron 74 y 70 características diferenciales, respectivamente. Además, la mayoría de las características enriquecidas en el grupo O30 generalmente exhibieron función de fermentación y capacidad de producir ácidos orgánicos, como ácidos grasos de cadena corta, ácido láctico y ácidos biliares, predominantemente en Bacteroidota y Firmicutes (Tabla S9) [47, 48, 49, 50, 51 , 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62] . Por ejemplo, Rikenella, Allobaculum, Bacteroides, Butyricicoccus, Faecalibaculum, f__Prevotellaceae, f__Ruminococcaceae, Lachnoclostridium, Lactobacillus y Parabacteroides
Entre las 50 características diferenciales correlacionadas positivamente con indicadores fisiológicos, 38 características exhibieron abundancias relativas más altas en el grupo O30 en comparación con los grupos C0 y S30. Estos taxones enriquecidos incluyeron Allobaculum, Candidatus_Saccharimonas, Dubosiella, Klebsiella, Lachnoclostridium, Lactobacillus, Muribaculum, Odoribacter, Parabacteroides, Prevotellaceae_UCG_001, Ruminococcaceae, entre otros. Además, la abundancia relativa de Alistipes, Bacteroides, Bilophila, Defluviimonas, Erysipelatoclostridium, Intestinimonas y Parasutterella fue mayor en el grupo O30 en comparación con el grupo C0 y se correlacionó positivamente con los indicadores fisiológicos. De manera similar, la abundancia relativa de Butyricicoccus, f__Prevotellaceae, Faecalibaculum, Fournierella y el grupo Lachnospiraceae NK4A136 se enriquecieron en el grupo O30 en comparación con el grupo S30, y también mostraron una correlación positiva con los indicadores fisiológicos (Figs. 7, S8).
Entre las 27 características diferenciales correlacionadas negativamente con indicadores fisiológicos, la abundancia relativa de 24 características fue significativamente menor en el grupo O30 en comparación con el grupo C0. Estas características incluyeron géneros como Aeromonas, Bryobacter, f__Methyloligellaceae, Motilimonas, Nautella, Pseudomonas, Sphingobacterium . Además, la abundancia relativa de Acinetobacter, Labilibacter y Nitrospira fue significativamen-
Fig 4. Análisis de diversidad alfa en los tipos de muestra de intestino, branquias, agua y WS. Se aplicó la normalización de la puntuación Z a cada índice de diversidad alfa. La agrupación en el mapa de calor se realizó mediante la distancia de Bray-Curtis con el método de ligamiento promedio. Las medias ± DE se muestran en la Tabla S 4. a, b, c Los valores que no comparten la misma letra en superíndice de cada columna son significativamente diferentes (P < 0,05). PD: diversidad filogenética; WS: fuente de agua. Información del grupo: C0: sin suplementación de Cu; S30: 30 mg/kg de suplementación de Cu inorgánico; O30: 30 mg/kg de suplementación de Cu orgánico.
te menor en el grupo O30 en comparación con el grupo S30, y estas características también mostraron una correlación negativa con los indicadores fisiológicos. Además, la abundancia relativa de Aeromonas y f__Chitinophagaceae fue notablemente menor en el grupo O30 en comparación con los grupos C0 y S30, y estas características se correlacionaron negativamente con los indicadores fisiológicos (Figs. 7, S8).
De las 36 funciones asociadas con características diferenciales, aquellas relacionadas con el ciclo del carbono (aeróbica-quimioheterotrofia, quimioheterotrofia y fermentación) se vincularon a un mayor número de características (Fig. 7). Además, las características enriquecidas en el grupo O30 o correlacionadas positivamente con indicadores fisiológicos pertenecían predominantemente a Bacteroidota y Firmicutes, con funciones principalmente asociadas con el ciclo del carbono y otros. Por el contrario, las características menos abundantes en el grupo O30 o correlacionadas negativamente con indicadores fisiológicos pertenecían predominantemente a Acidobacteriota, Actinobacteriota y Proteobacteria, con funciones anotadas principalmente relacionadas con los ciclos del carbono y el nitrógeno. Las características asociadas al ciclo del azufre incluyeron Salinisphaera (oxidación oscura de compuestos de azufre y oxidación oscura de tiosulfato) y Desulfovibrio (oxidación oscura de hidrógeno, respiración de compuestos de azufre y respiración de sulfato).
Las características clave y diferenciales se representaron con mayor detalle mediante diagramas de caja en las figuras S9 y S10, respectivamente. La composición de las características clave y diferenciales mostró una marcada distinción, con Nautella como característica clave únicamente en el grupo S30 y una característica diferencial enriquecida en el grupo C0. Además, la frecuencia general y la abundancia relativa de las características clave fueron mayores en todos los grupos (figura S9), mientras que la abundancia relativa de las características diferenciales se mantuvo generalmente por debajo del 1 % (figura S10).
Abundancia funcional de FAPROTAX Un total de 56 funciones ecológicamente relevantes fueron anotadas a través de la tabla ASV, las cuales fueron categorizadas en 6 grupos principales: ciclo del carbono, ciclo del nitrógeno, ciclo del azufre, ciclo del hierro, ciclo del manganeso, y otros (Fig. 8). En general, la abundancia de funciones relacionadas con los ciclos del carbono y nitrógeno fue mayor que otras funciones. Además, el grupo O30 exhibió una abundancia funcional mayor que los grupos C0 y S30 (Fig. 8 A). En las
Fig. 5. Gráficos de análisis de coordenadas principales (PCoA) para todos los tipos de muestra (A) y árboles de agrupamiento UPGMA (B) basados en distancias UniFrac no ponderadas para todos los grupos. Los valores de R² y P fueron los resultados de la prueba MANOVA permutacional general. WS: fuente de agua. Los grupos de muestra se designaron mediante prefijos para indicar los tipos de muestra: «I» para muestras intestinales, «G» para muestras de branquias y «W» para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
funciones relacionadas con el ciclo del carbono, la quimioheterotrofia aeróbica, la quimioheterotrofia y la fermentación fueron dominantes. La abundancia funcional de la fermentación fue mayor en el grupo O30 (27.7%) que en el grupo S30 (23.7%), que a su vez fue mayor que en el grupo C0 (19.9%). Las abundancias funcionales de la quimioheterotrofia aeróbica y la quimioheterotrofia fueron similares en los grupos S30 y O30, siendo ambas mayores que en el grupo C0. La abundancia funcional de la reducción de nitrato dominó el ciclo del nitrógeno. Fue mayor en el grupo O30 (26.9%) que en el grupo S30 (20.4%), que a su vez fue mayor que en el grupo C0 (18.6%). Además, las abundancias funcionales de desnitrificación, desnitrificación de nitrato, respiración de nitrato, desnitrificación de nitrito, respiración de nitrito, respiración de nitrógeno y desnitrificación de óxido nitroso fueron similares en los grupos C0 y O30,
ambas mayores que en el grupo S30, y la abundancia de estas funciones fue mayor al 1% en cada tratamiento. Otras abundancias funcionales también fueron generalmente mayores en el grupo O30, como la oxidación oscura de sulfuros, la oxidación oscura de compuestos de azufre, la respiración de hierro y manganeso, y otras (Fig. 8 B). El análisis PCA mostró que los grupos C0 y S30 tenían composiciones funcionales similares. Por el contrario, el grupo O30 formó un grupo distinto separado de los demás (Fig. 8 C).
Mapa de calor de la abundancia de funciones ecológicas potenciales basado en la base de datos FAPROTAX. A: Resumen de funciones anotadas. B: Funciones anotadas. Los números en las celdas de A y B representan la abundancia relativa promedio (%) de las características con funciones relevantes. Se aplicó la normalización de la puntuación Z a cada índice. C: Análisis de componentes
principales (PCA) basado en la composición de abundancia funcional mostrada en B. “−” indica que la característica no existía en ese grupo. Los grupos de muestra se designaron mediante prefijos para indicar los tipos de muestra: “I” para muestras intestinales, “G” para muestras de branquias y “W” para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
Predicción de funciones potenciales de Tax4Fun2
En la vía KEGG de nivel 1, se identificaron un total de 28 vías diferenciales, que pertenecen a 12 vías de nivel 2 (como transporte de membrana, metabolismo de lípidos y sistema digestivo) y 3 vías de nivel 3 (procesamiento de información ambiental, metabolismo y sistemas organismales) (Tabla S10).
En el intestino (Fig. 9 A), siete vías mostraron un enriquecimiento significativo en el grupo IO30 en comparación con los grupos IC0 y/o IS30, incluyendo el transportador ABC, la secreción biliar, la digestión y absorción de carbohidratos, entre otras. Además, el grupo IC0 presentó una abundancia significativamente menor del transportador ABC que el grupo IS30. Por el contrario, las vías metabólicas del ácido α-linolénico y de la tiamina se enriquecieron significativamente en el grupo IC0 en comparación con el grupo IS30.
Diagrama de cajas de las vías KEGG diferenciales de interés entre los grupos. a, b, c Las cajas de valores que no comparten la misma letra en superíndice son significativamente diferentes (Q < 0,25). Los grupos de muestra se designaron mediante prefijos para indicar los tipos de muestra: «I» para muestras intestinales, «G» para muestras branquiales y «W» para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
En la branquia (Fig. 9 B), se identificaron 19 vías diferenciales. Entre ellas, la vía de señalización cGMP-PKG, la biosíntesis de neomicina, kanamicina y gentamicina,
Fig 6. Análisis de la red de coocurrencia de la microbiota de las dietas C0, S30 y O30. Cada grupo de dieta contenía los tipos de muestra de intestino, branquias y agua. La agrupación de A y B se realizó utilizando la distancia de Bray-Curtis con el método de ligamiento promedio basado en los parámetros de las redes de coocurrencia microbiana proporcionados en la Tabla 2. A: nodos coloreados por la taxonomía del filo. B: nodos coloreados por la taxonomía de amplitud de nicho de la comunidad microbiana (NBMC). Las etiquetas representan las características clave; los colores de las etiquetas representan la NBMC de las características clave. C: gráfico de PCA basado en la NBMC de los nodos. D: diagrama de Sankey: características clave y sus funciones ecológicas basadas en la base de datos FAPROTAX. c__, clase y f__, familia. E: conectividad natural de la red. Los números dentro del gráfico representan la proporción de disminución de la conectividad. Los grupos de muestra se designaron mediante prefijos para indicar los tipos de muestra: «I» para muestras intestinales, «G» para muestras branquiales y «W» para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (30 mg/kg de suplementación de Cu inorgánico) y O30 (30 mg/kg de suplementación de Cu orgánico).
y la biosíntesis de estreptomicina se enriquecieron significativamente en el grupo GO30 en comparación con los grupos GC0 y GS30. Además, 9 vías, como el metabolismo de éter lipídico, la degradación de glicosaminoglicanos y la vía de señalización de la fosfolipasa D, se enriquecieron en los grupos GS30 y GO30 en comparación con el grupo GC0. Por el contrario, 7 vías, incluyendo la elongación de ácidos grasos, la absorción de minerales, el sistema de señalización de fosfatidilinositol y el metabolismo del azufre, exhibieron menor abundancia en el grupo GS30 en relación con los grupos GC0 y GO30. En el agua (Fig. 9 C), la abundancia de cinco vías fue significativamente mayor en los grupos WS30 y WO30 que en el grupo WC0, como la vía de señalización del calcio y la vía de señalización cGMPPKG. Además, la abundancia de tres vías fue significativamente mayor en el grupo WO30 que en el grupo WC0 o WS30, como el metabolismo del ácido dibásico ramificado en C5 y el sistema de señalización del fosfatidilinositol.
La presencia de características clave y diferenciales en los feeds
Un total de 36 características que estaban presentes tanto en las muestras de alimento como entre las características clave y diferenciales previamente identificadas (Tabla S11). El análisis de sus abundancias relativas reveló tres patrones distintos: Primero, Acinetobacter, Staphylococcus, Sphingobacterium y Bacillus mostraron abundancias relativas altas (> 2.0%) en las muestras de alimento, excediendo sus niveles en la microbiota del huésped. Segundo, se observaron niveles de abundancia moderados (0.5–1.0%) para Brevundimonas, Aeromonas y Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium, que aún eran más altos que los niveles del huésped. Tercero, Vibrio, Muribaculaceae, Pseudomonas y Lactobacillus mostraron abundancias relativas más bajas (0.1–0.5%) en el alimento en comparación con las muestras del huésped, con Vibrio mostrando una diferencia particularmente marcada. Los taxones restantes ocurrieron en niveles traza (< 0.05%), comparables a su abundancia en la microbiota del huésped.
Discusión
Cobre orgánico que promueve la utilización de nutrientes
La eficacia superior del cobre orgánico se debe principalmente a su capacidad única para prevenir la quelación de Cu 2+ por factores antinutricionales en fuentes de proteínas vegetales y para mitigar el antagonismo de minerales traza, como el antagonismo Zn-Cu [63]. Esta biodisponibilidad mejorada permite una utilización más eficiente del cobre por el organismo huésped [64]. Nuestros hallazgos experimentales corroboran estas ventajas, demostrando una acumulación y un ADC significativamente mayores tanto de Cu como de Zn en el grupo suplementado con cobre orgánico en comparación con los grupos de cobre inorgánico y control. Además, el grupo de cobre orgánico exhibió una acumulación mejorada de proteína cruda, carbono y nitrógeno, lo que sugiere una mejor asimilación de nutrientes y eficiencia metabólica. Estos hallazgos se alinean con y amplían nuestra investigación previa que demuestra los efectos beneficiosos del cobre orgánico en la salud fisiológica del camarón blanco [11].
El cobre orgánico modula la microbiota del camarón
Las composiciones de la dieta pueden afectar la microbiota del huésped. Aunque las diferencias en las dietas en este estudio fueron mínimas, variando solo en las fuentes de cobre, los resultados del análisis de conglomerados y la red de coocurrencia microbiana demostraron diferencias en la microbiota (intestino, branquias y agua). Específicamente, la red de coocurrencia
del grupo de cobre orgánico exhibió valores más altos de enlaces, grado promedio y modularidad relativa, junto con una longitud y diámetro de ruta promedio más bajos, lo que indica interacciones microbianas más fuertes. Combinados con una mayor conectividad natural y robustez, estos resultados sugirieron una red microbiana más estable y compleja en el grupo de cobre orgánico, que puede ayudar a los camarones a resistir estresores ambientales e invasiones de patógenos [65, 66]. Además, los cambios observados en la comunidad de microbiota se asociaron consistentemente con mejores indicadores fisiológicos en los camarones, lo que sugiere una modulación beneficiosa por el cobre orgánico. Estos hallazgos corroboran nuestras observaciones previas de mejores tasas de supervivencia bajo estrés de baja temperatura con suplementación de cobre orgánico [11]. Esto también indica que la presencia de camarones sanos y una red de coocurrencia estable se refuerzan mutuamente.
Las características clave (taxones clave) se consideran componentes fundamentales para mantener la estabilidad de la red de coocurrencia microbiana [65]. En comparación con las branquias y el agua circundante, las características clave del intestino son más significativas para proporcionar nutrientes esenciales y facilitar la absorción de nutrientes [67]. Casualmente, en este estudio, el grupo O30 mostró un conjunto único de características clave, incluyendo Candidatus_
Tabla 2. Parámetros de las redes de coocurrencia microbiana
Bacilloplasma, f__Mycoplasmataceae, Photobacterium y Vibrio, que generalmente se encuentran entre los 10 géneros principales en el intestino. Por el contrario, los grupos C0 y S30 compartieron características clave similares, incluyendo f__Arcobacteraceae, Leucothrix, Pseudarcobacter y Thiothrix, que se clasificaron predominantemente entre los 10 géneros principales en muestras de branquias y agua.
La investigación sobre el camarón blanco ha indicado que la abundancia de Candidatus_ Bacilloplasma generalmente se correlaciona positivamente con el estado de salud del camarón y es menor en el camarón enfermo [68, 69]. Además, las especies de Vibrionaceae (Vibrio y Photobacterium) generalmente se consideran endógenas y no patógenas [70]. Por lo tanto, como bacterias simbióticas en el intestino del camarón blanco, Candidatus_Bacilloplasma, Vibrio y Photobacterium, debido a sus posiciones clave en la estructura de la comunidad del grupo O30, pueden desempeñar papeles en la quimioheterotrofía, la fermentación y la inhibición de la proliferación de patógenos [71, 72]. Estos hallazgos sugirieron que el cobre orgánico puede ofrecer un beneficio superior en la mejora de la estructura de la microbiota del camarón. Además, en el presente estudio, las características clave de cada tratamiento no fueron necesariamente las más abundantes (Fig. S 9), lo que indica que el papel microecológico de un taxón dentro de la red microbiana es más crucial que su abundancia individual.
A diferencia de las características clave, las características diferenciales asociadas con las dietas y los indicadores fisiológicos fueron principalmente no dominantes (Fig. S10). A pesar de su baja abundancia, estas características pueden ejercer su influencia en el huésped, particularmente en el sistema inmunitario, a través del “efecto mariposa” o “efecto sinérgico” [73, 74, 75]. En particular, el grupo O30 exhibió un enriquecimiento de características productoras de ácidos orgánicos con posibles propiedades promotoras de la salud (Tabla S9). Se ha demostrado que los ácidos grasos de cadena corta y el ácido láctico mejoran el metabolismo energético, la respuesta inmunitaria y la barrera mucosa intestinal, así como que ejercen efectos antiinflamatorios [76]. Los ácidos biliares participan en varios procesos, incluida la
Fig. 7. Características de asociación significativas entre taxones microbianos y dietas, indicadores fisiológicos y posibles funciones ecológicas de estas características (base de datos FAPROTAX). Las leyendas de color se calcularon utilizando − log(valor q) × signo(coeficiente). Las celdas que indican asociaciones significativas están coloreadas (rojo o azul) y superpuestas con símbolos + o − para indicar una abundancia relativa mayor (+) o menor (−) de taxones microbianos al comparar dos dietas (las 3 columnas de la izquierda, las referencias de las dietas, son marrones), o una correlación positiva (+) o negativa (−) entre la abundancia relativa de taxones microbianos e indicadores fisiológicos. E_, Eubacterium. Los taxones no asignados a nivel de género están precedidos por una letra (o__, orden; f__, familia) que indica su nivel asignado. Información del grupo: C0, sin suplementación de Cu; S30, 30 mg/kg de suplementación de Cu inorgánico; O30, 30 mg/kg de suplementación de Cu orgánico.
desintoxicación, la función inmunitaria y la regulación de la homeostasis de la glucosa y el metabolismo lipídico [50]. Además, algunas características enriquecidas en el grupo O30 son resistentes a patógenos (f__Alteromonadaceae, Defluviimonas) [77, 78], o están involucradas en el ciclo del carbono (Thalassotalea) [79], nitrógeno (Klebsiella) [80], y azufre (Bilophila, Desulfovibrio, f__ Desulfovibrionaceae) [81]. Por el contrario, las características diferenciales que fueron
menos abundantes y correlacionadas negativamente con indicadores fisiológicos en el grupo O30 se identificaron predominantemente como Proteobacteria
Estas características son típicamente abundantes en camarones o peces enfermos, incluyendo Acinetobacter, Aquimarina, Brevundimonas, Dyella, Legionella y Nautella [82, 83, 84, 85, 86, 87]. Las diferencias en estas características también corrobora-
ron la promoción de la abundancia funcional de la microbiota en el grupo O30. Específicamente, la abundancia del ciclo del carbono (especialmente la fermentación), el ciclo del nitrógeno (especialmente la reducción de nitratos) y el ciclo del azufre (Fig. 6), y las vías KEGG relacionadas con el nivel 2 (p. ej., metabolismo de carbohidratos, metabolismo energético, metabolismo lipídico y sistema digestivo) (Tabla S10) fueron mayores en el grupo O30 que en los grupos C0 y S30. Los resultados demostraron que, además de consolidar los nichos microecológicos de las características dominantes, el cobre orgánico favoreció la regulación de la composición y la función de las características no dominantes dentro de la microbiota del camarón. Esto resultó en una microbiota más propicia para la salud del huésped, lo que coincidió con los hallazgos de los indicadores fisiológicos más saludables observados en el camarón alimentado con una dieta con cobre orgánico.
En este experimento, los nutrientes primarios en las dietas de camarones se mantuvieron en niveles constantes en todos los grupos. Sin embargo, la distinción en las fuentes de cobre, con 30 mg/kg, condujo a alteraciones notables en la microbiota del huésped, particularmente en el caso del cobre orgánico. Se observó que el cobre orgánico mejora las funciones ecológicas (principalmente los ciclos del carbono y el nitrógeno) y metabólicas. Estos hallazgos subrayaron la influencia de los micronutrientes, incluso en cantidades traza, y reforzaron la noción de que la microbiota desempeña un papel fundamental en el metabolismo de nutrientes del huésped. Además, sugirieron que las contribuciones microbianas al metabolismo de nutrientes fueron el resultado de los efectos sinérgicos de una amplia gama de miembros de la microbiota, incluyendo tanto bacterias dominantes como de baja abundancia [88].
La investigación sobre las interacciones entre el medio ambiente y la microbiota del huésped se ha centrado principalmente en cómo la calidad del agua impacta las comunidades de microbiota [89]. Sin embargo, nuestro estudio revela que la composición del alimento también puede influir en la composición y función de la microbiota acuática. Este fenómeno puede
Fig. 8. Mapa de calor de la abundancia de funciones ecológicas potenciales basado en la base de datos FAPROTAX.
A: resumen de las funciones anotadas. B: funciones anotadas. Los números en las celdas de A y B representan la abundancia relativa promedio (%) de las características con funciones relevantes. Se aplicó una normalización mediante Z-score a cada índice. C: análisis de componentes principales (PCA) basado en la composición de la abundancia funcional mostrada en B. El símbolo ‘–’ indica que la característica no estuvo presente en ese grupo.
Los grupos de muestras se designaron mediante prefijos que indican el tipo de muestra: ‘I’ para muestras intestinales, ‘G’ para muestras de branquias y ‘W’ para muestras de agua. Los grupos de tratamiento consistieron en C0 (control, sin suplementación de Cu), S30 (suplementación con Cu inorgánico a 30 mg/kg) y O30 (suplementación con Cu orgánico a 30 mg/kg).
estar asociado con una función metabólica mejorada y una microbiota intestinal modificada en el grupo de cobre orgánico del camarón. Por ejemplo, la composición de la microbiota acuática puede ser influenciada por la microbiota intestinal a través de la excreción fecal. Las altas tasas metabólicas pueden prevenir la acumulación de materia orgánica en el agua de cultivo, lo que también podría tener un efecto positivo en la composición de la microbiota acuática. Específicamente, observamos una acumulación y ciclos sustancialmente mayores de carbono y nitrógeno en el camarón. Esta mejora en los ciclos de carbono y nitrógeno puede impulsar el metabolismo energético del camarón.
El cobre desempeña un papel doble en las interacciones entre el camarón y la microbiota como un oligoelemento esencial y un agente antibacteriano [90, 91]. Las propiedades distintivas del cobre orgánico parecieron amplificar estos efectos beneficiosos. Los camarones del grupo del cobre orgánico exhibieron un enriquecimiento de una variedad diversa de bacterias que ejercen efectos beneficiosos sobre el huésped, acompañado de una abundancia relativa concurrente menor de ciertos patógenos potenciales. Sin embargo, se requiere más investigación para dilucidar los mecanismos por los cuales el huésped puede inhibir selectivamente a los patógenos sin alterar a los miembros beneficiosos de la microbiota. Es posible que, a lo largo del tiempo evolutivo, haya surgido una relación simbiótica entre el huésped y la microbiota. Esta relación puede permitir que el huésped utilice el cobre de manera estratégica, atacando a los patógenos y preservando a las bacterias críticas para la salud del huésped.
La selección del sitio corporal y del hospedador moldea la microbiota del camarón
La caracterización integral de la microbiota en múltiples nichos, incluidos el intestino, las branquias, el agua circundante y la fuente de agua, sigue siendo escasa en la investigación en acuicultura. Nuestro estudio destaca el fuerte impulso del sitio del cuerpo en la comunidad de microbiota en camarones (cultivados): Vibrio, Candidatus_Bacilloplasma y Photobacterium dominaron el intestino, Thiotrichaceae, c__Gammaproteobacte-
Fig. 9 The boxplot of differential KEGG pathways of interest among groups. a,b,c Value boxes not sharing the same superscript letter are significantly different (Q < 0.25). Sample groups were designated using prefixes to denote sample types: ‘I’ for intestinal samples, ‘G’ for gill samples, and ‘W’ for water samples. Treatment groups consisted of C0 (control, no Cu supplementation), S30 (30 mg/kg inorganic Cu supplementation), and O30 (30 mg/kg organic Cu supplementation).
ria y f__Saprospiraceae prevalecieron en las branquias, mientras que la microbiota en el agua circundante se distribuyó de manera relativamente uniforme. De manera similar, un estudio de 101 peces marinos (salvajes) reveló que el sitio del cuerpo ejerció la mayor influencia en la composición de la microbiota, superando tanto a las especies como a los factores del hábitat [12]. Además, una investigación de 89 peces salvajes reveló que la composición de la microbiota intestinal dependía del hábitat en lugar de estar determinada genéticamente [14]. Sin embargo, en los camarones cultivados, la tendencia observada pareció invertirse. Incluso en diferentes hábitats, se ha encontrado que Vibrio, Candidatus_Bacillo-
plasma y Photobacterium predominan de manera consistente en el intestino del camarón [9, 10, 69, 70, 92, 93, 94]. Además, desde la composición de la microbiota en la fuente de agua hasta el agua circundante, las branquias y, finalmente, el intestino, observamos un predominio gradual de taxones microbianos específicos, lo que indica una presión selectiva del huésped en la composición de la microbiota. Entre estos, las branquias parecían servir como un “puente” de transición que facilitaba la composición final de la microbiota intestinal dentro del camarón. El número de ASV compartidos entre los tipos de muestra siguió la tendencia: agua ∩ branquias > branquias ∩ intestino > intestino ∩ agua (Fig. S11). Además, los índices de di-
versidad alfa (ASV observados y PD de Faith) siguieron la tendencia: agua > branquias > intestino. Estos hallazgos respaldan el papel intermediario de las branquias. Los efectos de la selección del sitio corporal y del hospedador también se vieron influenciados por la adición de cobre en la dieta. En el grupo O30, las características principales (Fig. 3A) y las estructuras de la microbiota del intestino, las branquias y el agua del camarón blanco fueron más singulares (Fig. 5B). Además, el cobre orgánico favorece que la microbiota intestinal ocupe una posición más central entre los sitios corporales de intestino, branquias y agua (Fig. 6D y Fig. S9). Por el contrario, en el grupo con cobre inorgánico o sin adición de cobre, se identificaron características clave en las branquias y el agua.
Microbiota alimentaria y secuencias contaminantes
Los géneros dominantes en camarones estaban presentes en baja abundancia o ausentes en el alimento (Fig. S6), lo que sugiere que el impacto de la microbiota del alimento en el análisis de la microbiota del huésped en este estudio fue mínimo. Sin embargo, algunos géneros con muy baja abundancia en el huésped podrían verse afectados por el alimento, particularmente Acinetobacter, Staphylococcus, Sphingobacterium, Bacillus, Brevundimonas, Aeromonas y Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium (Tabla S11). Estos géneros se encuentran comúnmente en varios entornos (por ejemplo, suelo, agua y rizosfera) [95, 96, 97]. Este ADN microbiano puede estar presente en el huésped durante el proceso de cultivo. También existe la posibilidad de que el propio camarón sea la fuente de estos géneros, como Acinetobacter, Bacillus y Aeromonas, que son ubicuos. En general, el análisis de la microbiota alimentaria es necesario en los estudios que examinan la microbiota del huésped en acuicultura, ya que facilita una interpretación más objetiva de los resultados. Sin embargo, aparte del presente estudio, la secuenciación de la microbiota alimentaria sigue restringida a la investigación con salmón del Atlántico [18, 19].
Como punto final, la secuenciación de alto rendimiento mejora nuestra capacidad para realizar investigaciones nutricionales en alimentos acuícolas. Sin embargo, la contaminación de secuencias por reactivos y el me-
dio ambiente es común, y la falla en filtrar secuencias de ADN contaminantes puede llevar a una distorsión de datos en el análisis [23, 98]. No obstante, hay una escasez de investigaciones en acuicultura que aborden la eliminación de secuencias contaminantes de ADN durante el procesamiento de datos. Hasta donde sabemos, el procesamiento de datos relevante se ha reportado principalmente en estudios de peces, como el salmón del Atlántico [18, 25, 99, 100, 101, 102] y el róbalo común (Centropomus undecimalis) [103], así como nuestro estudio previo en camarón blanco [10]. En este estudio, también empleamos un proceso de eliminación de secuencias contaminantes. Encontramos que los resultados de las secuencias contaminantes fueron diferentes de nuestros resultados previos en camarón blanco, donde las principales secuencias contaminantes fueron o__Enterobacterales, Bryobacter, Blautia, Clostridium_sensu_stricto_7, Bacillus, entre otros [10]. También se ha observado variedad en secuencias contaminantes en estudios de salmón del Atlántico, pero se notó una mayor frecuencia de secuencias contaminantes de Pseudomonas en diferentes estudios [18, 25, 100]. Estas secuencias de Pseudomonas fueron consideradas a partir de la contaminación de reactivos [23, 98]. Sin embargo, en nuestro estudio, solo se identificó 1 ASV como Pseudomonas en la secuencia contaminante y en una abundancia muy baja (Tabla S5). Estos resultados resaltan la variabilidad de la contaminación en la secuenciación del gen 16s
rRNA, enfatizando la necesidad de remover secuencias contaminantes, ya que su presencia puede diferir considerablemente entre lotes de investigación y puede interferir con los estudios nutricionales en acuicultura.
Conclusiones
Los hallazgos actuales demostraron que el cobre orgánico regulaba los nichos microecológicos y mejoraba la estabilidad y la abundancia funcional de las comunidades de microbiota en el intestino, las branquias y el agua circundante. Este fenómeno se sinergizó con indicadores fisiológicos, promoviendo la salud general del camarón. Además, descubrimos que el cobre orgánico promovía la acumulación y los ciclos de carbono y nitrógeno en el camarón. En conclusión, nuestro estudio proporcionó una evaluación integral de las posibles aplicaciones del cobre orgánico para el cultivo saludable del camarón, ofreciendo nuevos conocimientos desde la perspectiva de la microbiota. Además, observamos las presiones selectivas impulsadas por el huésped que configuran la microbiota en diferentes sitios corporales y el papel de transición de las branquias en la configuración de la microbiota intestinal. Asimismo, nuestros hallazgos presentaron el primer análisis del impacto de las secuencias contaminantes y transmitidas por el alimento en los estudios de la microbiota del camarón, empleando métodos estadísticos para disminuir estas alteraciones•
Mioinositol oxigenasa: un actor clave en la lucha del camarón
blanco del Pacífico (Penaeus vannamei) contra el estrés por baja salinidad
La salinidad es un factor importante que afecta el entorno de producción de los animales acuáticos. La salinidad de un entorno puede incluso afectar directamente el crecimiento y la supervivencia de los organismos acuáticos (Bœuf y Payan, 2001). Cuando la salinidad cambia, los animales acuáticos ajustan su equilibrio de presión osmótica a través de órganos de ajuste osmótico y varios mecanismos de ajuste osmótico (Edwards y Marshall, 2012, Romano y Zeng, 2012, Takvam et al., 2021). Cuando la salinidad aumenta más allá del rango de tolerancia, el agua puede entrar y salir de las células a través de la ósmosis, causando deshidratación o hinchazón celular y, en última instancia, dando lugar a una serie de consecuencias fisiológicas, como daño celular y anomalías metabólicas (Kültz, 2015). Además, los cambios de salinidad pueden dañar a los animales acuáticos a través del estrés oxidativo, estrés del retículo endoplasmático, inflamación y muerte celular (Moniruzzaman et al., 2022, Paital y Chainy, 2010 ; Y. Wang et al., 2023). Por lo tanto, explorar el mecanismo regulador osmótico es la clave para abordar el estrés por salinidad. El mecanismo regulador osmótico es el proceso por el cual los organismos mantienen el equilibrio de la presión osmótica celular regulando las concentraciones de agua y solutos en el cuerpo. Con el desarrollo de la biología y la biotecnología, la investigación sobre la regulación osmótica ha logrado avances importantes, especialmente en las vías metabólicas, las vías de señalización y la regulación de la expresión génica (C. Su et al., 2023 ; X. Wang et al., 2023 ; Y. Wang et al., 2023). El estudio de los mecanismos reguladores osmóticos cubre varios aspectos, desde las vías metabólicas hasta la transducción de señales. Estudios han demostrado que diferentes organismos regulan la presión osmótica mediante complejas vías metabólicas y de señalización para proteger las células del estrés ambiental. Por lo tanto, estudios futuros revelarán con más detalle estos mecanismos y explorarán sus aplicaciones en la agricultura y la biotecnología.
El mioinositol es un alcohol de azúcar cíclico que está ampliamente presente en células animales y vegetales y es una molécula importante en muchos procesos
fisiológicos. El mioinositol desempeña múltiples funciones importantes en el metabolismo celular y la transducción de señales. No solo es un componente importante de la membrana celular, sino que también participa en la regulación de los niveles de iones intracelulares, el equilibrio metabólico , la proliferación celular y la apoptosis (Cui et al., 2022).
El estudio de las vías metabólicas del mioinositol es muy importante para comprender muchos procesos fisiológicos y mecanismos de regulación osmótica y proporciona objetivos potenciales para la acuicultura. La vía metabólica del mioinositol desempeña un papel crucial en las respuestas celulares a los desafíos de la presión osmótica (Alford et al., 2012). El metabolismo del mioinositol está regulado principalmente por la mioinositol oxigenasa (miox), y sus metabolitos pueden servir como sustratos para la síntesis de fosfatidilinositol (XB Su et al., 2023). El fosfatidilinositol, un componente crucial de la membrana celular, desempeña un papel vital en la transducción de señales durante el estrés osmótico (Pan et al., 2023). La investigación sobre Oreochromis niloticus reveló que el miox regula el equilibrio redox celular, ayuda en la adaptación al estrés abiótico y mejora la tolerancia a la sal en esta especie (Zhang et al., 2024). Al mismo tiempo, el miox es importante para mantener estables los niveles de mioinositol en el cuerpo. La investigación sobre cómo Penaeus vannamei responde al estrés de baja salinidad reveló que una cantidad apropiada de mioinositol puede mejorar la función antioxidante (Li et al., 2024).
El gen miox puede regular el nivel de mioinositol metabolizándolo, lo que afecta varios procesos fisiológicos en las células (Siddique et al., 2014). Numerosos estudios han demostrado que las concentraciones adecuadas de mioinositol no solo pueden mejorar el metabolismo de la glucosa y los lípidos y el rendimiento del crecimiento, sino que también ayudan a los animales acuáticos a afrontar mejor los desafíos osmóticos (Zhu et al., 2022, Chen et al., 2018). El metabolismo del mioinositol es crucial para la adaptación de los animales acuáticos a los cambios de salinidad (Kalujnaia et al., 2016, Ma et al.,
2020). Investigaciones preliminares han explorado el metabolismo del mio -inositol en peces para la adaptación a la salinidad; sin embargo, el papel del miox en crustáceos aún no se ha documentado.
El camarón blanco del Pacífico (P. vannamei) es una de las especies de acuicultura de mayor valor comercial en China y tiene muchas ventajas, como un alto rendimiento de carne, una rápida tasa de crecimiento y una fuerte adaptabilidad (Jamal, 2023, Marimuthu et al., 2024 ; Mustafa et al., 2023). Debido a su capacidad para vivir tanto en agua dulce como salada, P. vannamei sirve como un ejemplo clásico para investigar la transducción de señales de adaptación a la salinidad en crustáceos (Ye et al., 2024).
Nuestro estudio inicial reveló que el mioinositol puede aliviar significativamente la respuesta al estrés de P. vannamei en condiciones de baja salinidad a través de la vía de señalización del fosfatidilinositol, que se activa mediante receptores acoplados a la proteína G (Li et al., 2024). El consumo adecuado de carbohidratos aumenta la producción endógena de mioinositol y mejora la regulación osmótica en P. vannamei en condiciones prolongadas de baja salinidad (Lin et al., 2024). Las investigaciones han revelado que el metabolismo del mioinositol es crucial para la adaptación a la salinidad en P. vannamei Es probable que Pv-miox sea crucial para la regulación osmótica y la respuesta al estrés en condiciones de baja salinidad. En este estudio, se clonó e identificó la secuencia completa de ADNc de Pv-miox. Se evaluó el nivel de expresión del ARNm de Pv-miox en varios tejidos mediante PCR qRT. Se empleó la tecnología de interferencia de ARN (ARNi) para investigar la función de Pv-miox en la respuesta a la baja salinidad, lo que ofrece un nuevo marco teórico para examinar la regulación osmótica de los crustáceos y las vías de señalización.
Materiales y métodos
Animales de experimentación
El experimento utilizó P. vannamei suministrado por Hainan Zhongzheng Aquatic Technology Co., Ltd., China. El peso corporal inicial de P. vannamei fue de 0,5 ± 0,1 g. P. vannamei se
mantuvo temporalmente en agua de mar aireada filtrada a una temperatura de 26-29 , pH 7,8-8,3, una concentración de oxígeno disuelto de 6,1-6,8 mg/L, una salinidad de 29-32 y un nivel de nitrógeno amoniacal inferior a 0,04 mg/L. Durante el período de crianza temporal, P. vannamei se alimentó con alimento compuesto comercial para camarones (alimento Hainan Haitong) cuatro veces al día. La cantidad diaria de alimento fue del 6% del peso corporal y se ajustó según las condiciones reales. Las heces de los camarones y el alimento no consumido se eliminaron diariamente, seguido de un reemplazo del 50% del agua de mar filtrada y aireada. Tras un mes de cultivo temporal, se seleccionaron 5 ± 0,5 g de P. vannamei para experimentos posteriores. El estudio recibió la aprobación del Comité de Ética y Bienestar Animal de la Universidad de Hainan, Haikou, China (HNUAUCC-2024-00129).
Clonación de genes y análisis bioinformático Los camarones se seleccionaron de estanques de retención temporales, se congelaron rápidamente con nitrógeno líquido y se almacenaron a -80 para su uso posterior. El ARN total de las muestras se extrajo de acuerdo con las instrucciones del kit de extracción de ARN (9109, Takara, Japón). La pureza y la concentración del ARN total se evaluaron mediante espectrofotometría A260/A280 (NanoDrop One C, Thermo, EE. UU.) y la integridad de las bandas de ARN se verificó mediante electroforesis en gel de agarosa al 1%.
La construcción de la biblioteca de ADNc se llevó a cabo de acuerdo con las instrucciones del kit de reactivos PrimeScript™ RT (RR047A, Takara, Japón). El CDS previsto del gen Pv-miox (XM_027353045.1) se obtuvo de la base de datos del NCBI (https://www. ncbi.nlm.nih.gov/), y se diseñaron cebadores específicos con cebadores Pick, como se muestra en la Tabla 1. La región CDS dePvmioxse amplificó y clonó de acuerdo con las instrucciones de PrimeSTAR® HS (Premix) (R040A, Takara, Japón) y el kit de clonación pEASY®-Blunt (CB501, TransGen Biotech, China).
Tabla 1. Secuencias de pares de cebadores utilizadas para qRT-PCR
miox = mio -inositol oxigenasa nka = Na + /K + -ATPasa; nhe = Na + /H + intercambiador r ; cam = calmodulina; ca = anhidrasa carbónica; impa1 = mio -inositol monofosfatasa; sod = Superoxidasa dismutasa; gpx = Glutatión peroxidasa; nrf = Factor nuclear eritroide 2 relacionado con el factor 2 (2); bip = Proteína de inmunoglobulina de unión; ire1 = Enzima 1 que requiere inositol; xbp1s = Forma de empalme de la proteína de unión a X-box 1; nf-kb = Factor nuclear kappa B; tnf-α = Factor de necrosis tumoral alfa; jnk = Quinasa N-terminal de Jun; cytc = Complejo de citocromo; casp-3 = proteinasa específica de cisteinil aspartato 3; casp-9 = proteinasa 9 específica de cisteinil aspartato.
El marco de lectura abierto (ORF) de la secuencia de nucleótidos de Pv-miox se predijo y se tradujo en aminoácidos mediante el buscador de ORF (https://www.ncbi.nlm.nih. gov/orffinder/). La secuencia de aminoácidos predicha se comparó con la base de datos de proteínas mediante la herramienta BLAST en NCBI (https://blast.ncbi.nlm.nih.gov/ Blast.cgi). El contenido de aminoácidos, el punto isoeléctrico predicho y la masa molecular teórica se analizaron mediante ExPASy ProtParam (https://web.expasy.org/ protparam/). El dominio estructural de la proteína de Pv-miox se examinó mediante SMART 4.0 (http://smart.embl-heidelberg. de/). Los sitios de glicosilación se predijeron mediante el servidor NetNGlyc 1.0 (http:// www.cbs.dtu.dk/services/NetNGlyc/). Los sitios de fosforilación se predijeron mediante el servidor NetPhos 3.1 (http:// www.cbs.dtu.dk/services/NetPhos/). La estructura tridimensional de la proteína se predijo mediante SWISS-MODEL (https:// swissmodel.expasy.org/). El método SOPMA se utiliza para predecir la estructura secundaria de una proteína. El árbol filogenético se desarrolló mediante los programas DNAMAN 9.0 y MEGA 11.0.
Expresión tisular diferencial
Se seleccionaron doce camarones intactos y sanos para la toma de muestras; tres de ellos se mezclaron en cada tubo y se realizaron cuatro réplicas biológicas. El orden de muestreo fue el siguiente: hemolinfa, branquias, pedúnculo ocular, hepatopáncreas, intestino, cerebro y músculo. Todos los tejidos se colocaron en crioviales, se congelaron rápidamente con nitrógeno líquido y se almacenaron a -80 °C hasta su posterior análisis de expresión diferencial.
Preparación de dsRNA y verificación de la eficacia de la interferencia
Diseño de cebadores para la síntesis de dsRNA basado en la secuencia de nucleótidos de Pv-miox clonada en este estudio. Se utilizó la herramienta en línea
SnapDragon (https://www.flyrnai.org/ snapdragon) para predecir secuencias diana con múltiples sitios de unión funcionales de ARN de interferencia para el diseño de cebadores de dsRNA de Pv-miox. Siguiendo las instrucciones del sistema T7 RiboMAX™ Express RNAi, se añadieron secuencias
del promotor T7 a ambos extremos de los cebadores y se sintetizó el dsRNA de Pvmiox siguiendo estos pasos. Además, se sintetizó dsRNA de EGFP como control.
Un total de 72 camarones sanos fueron asignados a 9 grupos, cada uno compuesto por 8 camarones. Los camarones recibieron inyecciones de ARNdc de Pv-miox, ARNdc de EGFP (control positivo) y PBS (control negativo) en concentraciones de 3, 5 y 7 µg/g, respectivamente, en función del peso corporal, y se administraron en los músculos del segmento de la cola del camarón. Se recolectaron muestras de branquias 24 h después de la inyección, se congelaron rápidamente en nitrógeno líquido y posteriormente se almacenaron a -80 para evaluar los efectos de las diferentes concentraciones. Se seleccionó un total de 72 camarones sanos y se dividieron en 3 grupos, con 24 camarones en cada grupo. Cada grupo se dividió en tres puntos de tiempo, con 8 camarones en cada punto de tiempo. A los tres grupos de camarones se les inyectó 7 μg/g (según el peso corporal) de ARNdc de Pv-miox, ARNdc de EGFP y PBS en el segundo segmento abdominal.
Se tomaron muestras de las branquias 24, 48 y 72 h después de la inyección. Las muestras se congelaron rápidamente con nitrógeno líquido y se almacenaron a -80 °C para determinar el tiempo efectivo de interferencia. Los cebadores utilizados para sintetizar el ARNdc se muestran en la Tabla 1.
Estadísticas de mortalidad y recolección de muestras tras la interferencia de ARN En este estudio, se investigó la mortalidad en condiciones agudas de baja salinidad después de la interferencia de ARN, y se establecieron tres grupos experimentales (n = 90 camarones/grupo) mediante aleatorización estratificada. Cada grupo contenía tres réplicas biológicamente independientes (30 camarones/réplica) alojadas en diferentes unidades de cultivo, asegurando un aislamiento físico completo entre las unidades experimentales.
Los ratones ayunaron durante 24 h antes del experimento. El segundo segmento abdominal se inyectó con ARNdc de Pv-miox, ARNdc de EGFP y PBS. Los camarones de los
tres grupos se sometieron a estrés de baja salinidad 24 h después de la inyección, y se registró la mortalidad a las 12, 24 y 48 h para el análisis acumulativo. Simultáneamente, se utilizaron 360 camarones para evaluar el impacto del estrés agudo de baja salinidad después de la interferencia de ARN. Los 360 camarones se asignaron a 3 grupos de 120 camarones cada uno, y cada grupo se subdividió en 4 puntos de tiempo, que comprendían 30 camarones por punto de tiempo. El segundo segmento abdominal se inyectó con ARNdc de Pv-miox, ARNdc de EGFP y PBS. Los camarones de los tres grupos se expusieron a estrés por baja salinidad 24 h después de la inyección. Se recolectaron tejidos de hemolinfa, branquias y hepatopáncreas a las 0, 12, 24 y 48 h, se congelaron rápidamente en nitrógeno líquido y se almacenaron a -80 °C para su posterior análisis.
Análisis bioquímico
La muestra de hemolinfa se mantuvo a 4 °C durante la noche, se descompuso la masa coagulada y se centrifugó a 3500 rpm durante 15 min. El sobrenadante resultante fue suero. Los niveles de Na +, K +, Cl₂ y Ca₂ en el suero se determinaron mediante kits de detección proporcionados por el Instituto de Bioingeniería Jiancheng de Nanjing. Los niveles de mioinositol en las branquias y el hepatopáncreas se determinaron mediante kits ELISA (Shanghai Enzyme Biotechnology Company, China).
PCR cuantitativa en tiempo real (qRT-PCR)
La secuencia del gen diana utilizada para la cuantificación de fluorescencia se obtuvo de NCBI (https://www.ncbi.nlm.nih.gov/), y los cebadores de cuantificación de fluorescencia se diseñaron utilizando Pick-Primers (Tabla 1). El gen diana se amplificó por qRTPCR utilizando el sistema LightCycler®96 (Roche, Suiza). El sistema qRT-PCR y los procedimientos se realizaron de acuerdo con las instrucciones del fabricante para ChamQ Universal SYBR qRT-PCR Master Mix (Q71102, Vazyme, China). Los genes de referencia dual utilizados fueron beta-actina (β-actina) y factor de elongación 1-alfa-similar (EF1-α). La estabilidad de los cebadores se evaluó a través de RefFinder (http://blooge. cn/RefFinder/). La especificidad de la amplificación se verificó mediante análisis de curva de fusión. La eficiencia de amplificación
de todos los cebadores estuvo entre el 90% y el 110%. La expresión relativa de los genes diana se calculó mediante el método 2 -ΔΔCt .
Análisis estadístico
Todos los análisis estadísticos se realizaron con SPSS Statistics 27 (IBM, Armonk, NY, EE. UU.). Todos los datos se distribuyeron normalmente y presentaron varianzas homogéneas. La significancia de las diferencias entre los diferentes grupos de tratamiento se analizó mediante un análisis de varianza unidireccional y la técnica de comparación múltiple de Duncan. Los resultados se expresan como media ± error estándar (media ± EE). Un valor de p < 0,05 indicó que la diferencia era estadísticamente significativa. Los resultados del análisis se visualizaron con GraphPad Prism 8. El análisis de correlación de Pearson se realizó con datos de expresión génica (https://www. chiplot.online/mantel_test_correlation_ heatmap.html).
Resultados
Análisis de secuencia y homología de Pvmiox
La región codificante de Pv-miox se amplificó y secuenció según la base de datos del genoma completo del NCBI. Los resultados de la secuenciación coincidieron con la secuencia génica prevista de Pv-miox de P. vannamei. La longitud total del ADNc de Pv-miox es de 1088 pb (fig. 1 A). El marco de lectura abierto (ORF) es de 879 pb y puede codificar 292 aminoácidos. ExPASy-ProtParam predijo que su peso molecular era de 34,12 kD y su punto isoeléctrico teórico era de 5,71. El índice de inestabilidad previsto (₂) era de 42,41, lo que indicaba que la proteína era inestable. La hidrofilia media (GRAVY) era de − 0,649 y el índice alifático era de 67,97. El análisis de predicción estructural reveló que Pv-miox tiene siete dominios funcionales. El programa SOPMA reveló que la proteína MIOX contenía un 46,39% de hélice α, un 5,5% de cadena extendida y un 48,11% de hélice aleatoria (Fig. 1 B y C). Pv-miox posee varios sitios funcionales. Según NetPhos
3.1, Pv-miox contiene 15 sitios de serina, 16 sitios de treonina y 17 sitios de tirosina, para un total de 48 sitios de fosforilación (Fig. 1 D).
La secuencia de aminoácidos de Pvmiox se analizó mediante BLAST en NCBI para compararla con secuencias de otras especies (Fig. 2 A). El estudio reveló que Pvmiox tiene una alta homología con el gen Pvmiox de Penaeus indicus, con una similitud del 95,2%. La secuencia de aminoácidos de Pv-miox se analizó junto con otras especies y se generó un árbol filogenético mediante el software DNAMAN 9.0 y MEGA 11.0. P. vannamei está resaltado en rojo (Fig. 2 B). El árbol filogenético se divide en tres taxones principales: crustáceos, insectos y peces. Los crustáceos están resaltados con un fondo verde, los insectos con un fondo azul y los peces con un fondo gris.
Expresión de Pv-miox en diferentes tejidos Se detectaron los niveles de expresión de Pvmiox en la hemolinfa, el pedúnculo ocular,
Fig. 1. Análisis de la secuencia de Pv-miox e ilustración de la proteína. (A) Secuencia de aminoácidos del gen Pv-miox y ADNc completo. El texto en rojo indica el codón de inicio (ATG) y el codón de terminación (TAA); los recuadros indican los dominios funcionales característicos de la anhidrasa carbónica, la glicosilhidrolasa y la fosfohidrolasa (y otros residuos de aminoácidos de moléculas pequeñas). (B) Estructura tridimensional (3D) de la proteína Pv-miox. (C) Análisis de la estructura secundaria : hélice α (azul), cadena extendida (morado) y espiral aleatoria (amarillo). (D) Sitios de fosforilación predichos.
Fig. 2. Alineamiento de secuencias múltiples y análisis del árbol filogenético. (A) Alineamiento múltiple de la secuencia de aminoácidos de Pv-miox en P. vannamei con la de otras especies. (B) Árbol filogenético NJ de Pv-miox con la de otras especies.
las branquias, el hepatopáncreas, el cerebro, el intestino y el músculo de P. vannamei mediante qRT-PCR (Fig. 3). Los resultados revelaron que Pv-miox se expresó en todos los tejidos de P. vannamei analizados, con el mayor contenido en el hepatopáncreas, seguido de las branquias, el músculo y el intestino, y el menor contenido en la hemolinfa, el cerebro y el pedúnculo ocular.
Eficiencia de interferencia y mortalidad
Entre los tejidos analizados, el nivel de expresión de Pv-miox fue mayor en el hepatopáncreas. Dado que las branquias se seleccionaron como el tejido principal para la detección en este estudio, también se utilizaron como objeto de evaluación para la eficiencia de la interferencia de ARN.
Se evaluó la eficiencia de interferencia del ARNdc de Pv-miox a dosis de 3 μg/g, 5 μg/g y 7 μg/g de peso corporal (Fig. 4 A). Los resultados indicaron que la dosis de 7 μg/g de peso corporal fue la más efectiva para la interferencia. La duración de la interferencia después de la inyección de ARNdc de Pv-miox fue de 7 μg/g de peso corporal (Fig. 4 B). Estos resultados indicaron que la eficiencia de interferencia se mantuvo durante 72 h.
Fig. 3. Expresión de Pv-miox en diferentes tejidos. Las barras de error indican ± EEM (n ≥ 3). Se utilizó el método Livak (2 -ΔΔCt) para normalizar el nivel de expresión del gen diana, utilizando β-actina y EF1α como genes de referencia internos. Las diferentes letras en superíndice (a, b, c, d) indican diferencias significativas entre los distintos grupos (p < 0,05).
El experimento de ARNi evaluó las tasas de mortalidad en varios grupos a distintos intervalos de tiempo tras la exposición al estrés por salinidad 3 (Fig. 4 C). El grupo de ARN bicatenario Pv-miox presentó sistemáticamente una tasa de mortalidad menor que la de los grupos de ARN bicatenario PBS y EGFP. Las tasas de mortalidad no
difirieron significativamente entre los grupos de ARN bicatenario PBS y EGFP.
Síntesis y metabolismo del mioinositol
La expresión del gen impa1 en cada grupo de tratamiento aumentó con el tiempo; sin embargo, no hubo diferencia significativa en la expresión del gen impa1 entre los grupos
PRODUCCIÓN
de tratamiento (Fig. 5 A). La expresión del gen Pv-miox en el grupo Pv-miox dsRNA fue significativamente menor que en el grupo PBS y el grupo EGFP dsRNA a las 0, 12 y 48 h, y la expresión del gen Pv-miox en cada grupo de tratamiento fue significativamente menor que a las 0 h (Fig. 5 B). La concentración de mioinositol en las branquias del grupo Pvmiox dsRNA fue significativamente mayor que en las branquias del grupo PBS y el grupo EGFP dsRNA a las 12 h, y no hubo diferencia significativa entre los grupos de tratamiento en otros puntos temporales (Fig. 5 C). La concentración de mioinositol en el hepatopáncreas del grupo de ARN bicatenario Pv-miox fue significativamente mayor que la del grupo de PBS y la del grupo de ARN bicatenario EGFP, y la concentración de mioinositol en el hepatopáncreas de cada grupo de tratamiento disminuyó significativamente con el tiempo (Fig. 5 D).
Concentración de iones en suero
Efectos de la interferencia de ARN en las concentraciones de Na +, K +, Cl - y Ca 2+ en suero de camarón bajo estrés agudo de baja salinidad (Fig. 6). El contenido de Na + en cada grupo de tratamiento tendió a disminuir con el tiempo, y el contenido de Na + en el grupo de ARN bicatenario Pv-miox fue significativamente mayor que en el grupo de PBS y el grupo de ARN bicatenario EGFP (Fig. 6 A). El contenido de K + en cada grupo de tratamiento tendió a disminuir primero pero luego a aumentar con el tiempo, y el contenido de K + en el grupo de tratamiento de ARN bicatenario Pv-miox fue significativamente mayor que en el grupo de PBS y el grupo de ARN bicatenario EGFP (Fig. 6 B). El contenido de Ca 2+ en el grupo de ARN bicatenario Pvmiox fue el más alto a las 12 h, mientras que el contenido de Ca 2+ en el grupo de PBS y el grupo de ARN bicatenario EGFP no cambió significativamente con el tiempo (Fig. 6 C). No hubo diferencia significativa en el contenido de Cl - entre los grupos de tratamiento (Fig. 6 D).
Expresión de ARNm de genes relacionados con el transportador de iones en branquias El impacto de la interferencia de ARN en los niveles de expresión de ARNm de nka, nhe, ca y cam en branquias sometidas a estrés agudo por baja salinidad (Fig. 7). La expresión del gen nka aumentó con el tiempo en todos los grupos de tratamiento,
Fig. 4. Comparación de diferentes eficiencias de interferencia de dsRNA y tasas de mortalidad. (A) Dosis de inyección; (B) Tiempo de inyección; (C) Mortalidad. Las barras de error indican ± EEM (n ≥ 3). Las letras en superíndice (a, b, c) indican diferencias significativas entre los distintos grupos (p < 0,05); “*” indica p ≤ 0,05.
Fig. 5. Efectos de la interferencia de dsRNA en la síntesis y metabolismo de mioinositol en P. vannamei bajo estrés agudo por baja salinidad. Las barras de error indican ± SEM (n ≥ 3). (A) impa1 en branquias; (B) Pv-miox en hepatopáncreas; (C) Contenido de mioinositol en branquias; (D) Contenido de mioinositol en hepatopáncreas. Se utilizó el método Livak (2 -ΔΔCt) para normalizar el nivel de expresión del gen diana, con β-actina y EF-1α utilizados como genes de referencia internos. Diferentes letras en superíndice (a, b, c) indican diferencias significativas entre diferentes grupos (P < 0,05). “**” indica P ≤ 0,01. impa1 = inositol monofosfatasa
y el grupo de ARNdc Pv-miox mostró una expresión significativamente mayor que los grupos de ARNdc PBS y EGFP (Fig. 7 A).
La expresión del gen nhe en el grupo de ARNdc Pv-miox se elevó significativamente en comparación con los grupos de ARNdc PBS y EGFP a las 12, 24 y 48 h, observándose el pico de expresión a las 24 h (Fig. 7 B).
La expresión del gen ca en el grupo de ARNdc Pv-miox se elevó significativamente en comparación con los grupos de ARNdc PBS y EGFP a las 0, 12 y 24 h, observándose la expresión más alta a las 24 h (Fig. 7 C).
La expresión del gen cam aumentó con el tiempo en todos los grupos de tratamiento, y el grupo de ARNdc Pv-miox mostró una expresión significativamente mayor que los grupos de ARNdc PBS y EGFP a las 0 y 12 h (Fig. 7 D).
Expresión de ARNm de genes relacionados con el estrés oxidativo y el estrés del retículo endoplasmático en branquias
La expresión del gen sod en el grupo Pvmiox dsRNA fue notablemente elevada en comparación con los grupos PBS y EGFP dsRNA a las 0, 12 y 24 h, observándose el pico de expresión a las 24 h (Fig. 8 A). La expresión del gen gpx en el grupo Pv-miox dsRNA fue significativamente elevada en comparación con los grupos PBS y EGFP dsRNA tanto a las 0 como a las 12 horas, observándose la expresión más alta a las 12 h (Fig. 8 B). La expresión del gen nrf2 en el grupo Pvmiox dsRNA fue significativamente elevada en comparación con los grupos PBS y EGFP dsRNA, observándose la expresión más alta a las 24 h (Fig. 8 C). Los niveles de expresión del gen bip inicialmente aumentaron y luego disminuyeron con el tiempo en todos los grupos de tratamiento, sin observarse diferencias significativas entre ellos (Fig. 8 D). La expresión del gen ire1 en el grupo de dsRNA Pv-miox se elevó significativamente a las 12, 24 y 48 h, y la expresión más alta se observó a las 12 h (Fig. 8 E). La expresión del gen xbp1s en el grupo de dsRNA Pv-miox se elevó significativamente en comparación con la de los grupos de dsRNA PBS y EGFP a las 0, 12 y 24 h, y la expresión más alta se observó a las 24 h (Fig. 8 F).
Expresión de ARNm de genes relacionados con la inflamación y la apoptosis en las branquias
Los niveles de expresión del gen nf- κ b en
Fig. 6. Efectos de la interferencia de dsRNA en el contenido iónico sérico de P. vannamei bajo estrés agudo por baja salinidad. Las barras de error indican ± EEM (n ≥ 3). (a) Na + ; (b) K + ; (c) Cl1 ; (d) Ca22. Las letras en superíndice (a, b, c) indican diferencias significativas entre los distintos grupos (p < 0,05). “*” indica p ≤ 0,05 y “**” indica p ≤ 0,01.
Fig. 7. Efectos de la interferencia de dsRNA en genes relacionados con transportadores de iones en las branquias de P. vannamei bajo estrés agudo de baja salinidad. Las barras de error indican ± SEM (n ≥ 3). (A) Na + /K + -ATPasa (nka); (B) Intercambiador Na + /H + (nhe); (C) anhidrasa carbónica (ca); (D) calmodulina (cam). El método Livak (2 -ΔΔCt) se utilizó para normalizar el nivel de expresión del gen diana, con β-actina y EF-1α utilizados como genes de referencia internos. Diferentes letras en superíndice (a–c) indican diferencias significativas entre diferentes grupos (P < 0,05). “*” indica P ≤ 0,05 y “**” indica P ≤ 0,01.
el grupo de ARN bicatenario Pv-miox fueron significativamente mayores que los del grupo PBS y el grupo de ARN bicatenario EGFP a las 12 y 24 h, y el nivel de expresión del gen nf - κ b en el grupo de ARN bicatenario Pv-miox fue el más alto a las 24 h (Fig. 9 A). Los niveles de expresión del gen tnf-α en el grupo de ARN bicatenario Pvmiox fueron significativamente mayores
que los del grupo PBS y el grupo de ARN bicatenario EGFP, y los niveles de expresión del gen tnf-α en cada grupo de tratamiento mostraron una tendencia creciente con el tiempo (Fig. 9 B). Los niveles de expresión del gen jnk en el grupo Pv-miox dsRNA fueron significativamente más altos que aquellos en el grupo PBS y el grupo EGFP dsRNA a las 0, 12 y 24 h, y el nivel de expresión del gen jnk en
el grupo Pv-miox dsRNA fue el más alto a las 24 h (Fig. 9 C). Los niveles de expresión del gen cytc en el grupo Pv-miox dsRNA a las 24 y 48 h fueron significativamente más bajos que aquellos en el grupo PBS y el grupo EGFP dsRNA, y los niveles de expresión del gen cytc en cada grupo de tratamiento aumentaron con el tiempo (Fig. 9 D). Los niveles de expresión del gen casp-3 en el grupo Pv-miox dsRNA a las 24 y 48 h fueron significativamente más bajos que aquellos en el grupo PBS y el grupo EGFP dsRNA, y los niveles de expresión del gen casp-3 en cada grupo de tratamiento aumentaron con el tiempo (Fig. 9 E). Los niveles de expresión del gen casp-9 en el grupo de dsRNA Pvmiox fueron significativamente inferiores a los del grupo PBS y el grupo de dsRNA EGFP a las 12, 24 y 48 h, y los niveles de expresión del gen casp-9 en cada grupo de tratamiento aumentaron con el tiempo (Fig. 9 F).
Correlación de indicadores fisiológicos
El análisis de correlación de Pearson (Fig. 10) mostró que el gen Pv-miox se correlacionó significativamente de forma positiva con cytc, casp-9, casp-3 y la mortalidad, y significativamente de forma negativa con Na +, Ca2 +, nka, nhe, sod, nrf2, ire1, nfkb, tnf-a y jnk. El gen impa1 se correlacionó significativamente de forma positiva con nhe, sod, nrf2, ire1, nf- κ b y jnk, y significativamente de forma negativa con cytc, casp-9, casp-3 y la mortalidad. El mioinositol se correlacionó significativamente de forma positiva con Na +, K +, nka, nhe, ca, sod, nrf2, ire1, nf- κ b y tnf-α y se correlacionó negativamente con casp-9 y casp-3
Discusión
En este estudio, el ADNc de Pv-miox de longitud completa se clonó de P. vannamei. El análisis de la secuencia de aminoácidos mostró que tiene 7 dominios transmembrana. Tiene el dominio característico de los dominios funcionales de anhidrasa carbónica , glicosil hidrolasa y fosfohidrolasa (y otros residuos de aminoácidos de molécula pequeña).
Estos dominios funcionales podrían ser cruciales para el metabolismo del mioinositol. El Pv-miox de P. vannamei estaba más estrechamente relacionado con el de Penaeus indicus, agrupándose en una rama (con una similitud de hasta el 95,2%). Además, P. vannamei y otros crustáceos
Fig. 8. Efectos de la interferencia de dsRNA en genes relacionados con el estrés oxidativo en las branquias de P. vannamei bajo estrés agudo de baja salinidad. Las barras de error indican ± SEM (n ≥ 3). (A) Superoxidasa dismutasa (sod); (B) Glutatión peroxidasa (gpx); (C) Factor nuclear eritroide 2 relacionado con el factor 2 (nrf2); (D) Proteína de inmunoglobulina de unión (bip); (E) Enzima 1 que requiere inositol (ire1); (F) Forma de empalme de la proteína de unión a X-box 1 (xbp1s). Se utilizó el método Livak (2 -ΔΔCt) para normalizar el nivel de expresión del gen diana, con β-actina y EF-1α utilizados como genes de referencia internos. Diferentes letras en superíndice (a–c) indican diferencias significativas entre diferentes grupos (P < 0,05). “*” indica P ≤ 0,05 y “**” indica P ≤ 0,01.
Fig. 9. Efectos de la interferencia de dsRNA en genes relacionados con la inflamación en las branquias de P. vannamei bajo estrés agudo de baja salinidad. Las barras de error indican ± SEM (n ≥ 3). (A) Factor nuclear kappa B (nf-kb); (B) Factor de necrosis tumoral alfa (tnf-α); (C) Jun N-terminal quinasa (jnk); (D) Complejo de citocromo (cytc); (E) proteinasa específica de cisteinil aspartato 3 (casp-3); (F) proteinasa específica de cisteinil aspartato 9 (casp-9). Se utilizó el método Livak (2 -ΔΔCt) para normalizar el nivel de expresión del gen diana, con β-actina y EF-1α utilizados como genes de referencia internos. Diferentes letras en superíndice (a–c) indican diferencias significativas entre diferentes grupos (P < 0,05). “*” indica P ≤ 0,05 y “**” indica P ≤ 0,01.
se agruparon en una rama amplia a una distancia evolutiva relativamente alta de los insectos, lo que indica que el Pvmiox estaba altamente conservado durante la evolución. La alta conservación evolutiva del Pv-miox indica que desempeña un papel fisiológico clave en múltiples especies,
especialmente en la regulación de la presión osmótica, el metabolismo energético y la respuesta al estrés ambiental. Por ejemplo, en un estudio de tilapia del Nilo, se demostró que MIOX desempeña un papel importante en la regulación osmótica y mejora la tolerancia a la sal de la tilapia del Nilo al
afectar la acumulación de inositol , el metabolismo de la glucosa y las propiedades antioxidantes (Zhang et al., 2024). Los resultados de la expresión diferencial del tejido revelaron que Pv-miox se expresó ampliamente en todos los tejidos probados. El nivel de expresión de Pv-miox en el hepatopáncreas del camarón fue el más alto. El hepatopáncreas es un tejido metabólico importante para el camarón (Chen et al., 2015). El exceso de mioinositol en el camarón puede transportarse al hepatopáncreas para el metabolismo, que desempeña un papel importante en el mantenimiento de la homeostasis del mioinositol en el cuerpo y el mantenimiento de varias funciones fisiológicas (Bu et al., 2021, Croze y Soulage, 2013, Shirmohammad et al., 2016).
Como regulador osmótico orgánico, el mioinositol desempeña un papel irremplazable en la respuesta del camarón a los cambios de salinidad (Chen et al., 2018). Muchos estudios han demostrado que añadir mioinositol dietético al alimento mejora significativamente la tasa de supervivencia, el rendimiento del crecimiento y la capacidad de regulación osmótica del camarón en condiciones de baja salinidad (Guilun et al., 2022, Lu et al., 2023, Yu et al., 2023). La función del mioinositol es inseparable del importante papel de la síntesis de mioinositol y las vías metabólicas. Muchos estudios han demostrado que los genes relacionados con la síntesis de mioinositol desempeñan papeles importantes en la regulación osmótica (Gardell et al., 2013, Hamar, 2024, Sacchi et al., 2014). Sin embargo, el papel del metabolismo del mioinositol en los crustáceos aún no está claro. Este estudio verificó la relación entre el metabolismo del mioinositol y la adaptación a la salinidad bajo estrés de baja salinidad a través de experimentos de interferencia de dsRNA. Los resultados de qRT-PCR mostraron que el grupo Pv-miox dsRNA presentó una expresión significativamente menor que el grupo control, lo que indica que la interferencia fue efectiva. La tasa de mortalidad después de la interferencia de dsRNA Pv-miox fue significativamente menor que en los otros grupos. Además, el nivel de expresión del gen impa1 de la biosíntesis de mioinositol aumentó y la concentración de mioinositol se acumuló
Fig. 10. Análisis de correlación de indicadores fisiológicos. Las líneas azul y roja representan correlaciones negativas y positivas, respectivamente; el ancho de la línea indica la magnitud del coeficiente de correlación de Mantel; la línea continua indica una correlación significativa (p < 0,05) y la línea punteada, una correlación insignificante (p ≥ 0,05). El gradiente de color de azul (correlación negativa) a rojo (correlación positiva) representa el coeficiente de correlación de Pearson.
en las branquias. La posible razón de este fenómeno es que cuando cambia la salinidad externa, la acumulación de mioinositol regula la actividad de los transportadores de iones, lo que ayuda a mantener el equilibrio de la presión osmótica de las células (Zhu et al., 2021, Gardell et al., 2013, Strange et al., 1991). Sin embargo, el metabolismo del mioinositol es muy importante para la homeostasis del mismo Los resultados mostraron que el contenido de mioinositol en el hepatopáncreas aumentó, lo que indica que su metabolismo se redujo, lo que confirma que el gen Pv -miox desempeña un papel clave en la adaptación del camarón a ambientes de baja salinidad. Por lo tanto, realizamos experimentos adicionales para verificar el papel del gen Pv-miox en la adaptación a baja salinidad.
Las branquias de P. vannamei son los principales órganos involucrados en el mantenimiento de la presión osmótica y la regulación iónica. Los transportadores de iones ubicados en el epitelio branquial desempeñan un papel vital en la regulación osmótica (Pan et al., 2014). Por ejemplo, cuando se enfrentan al estrés de baja salinidad, la Na + /K + -ATPasa en las branquias promueve la absorción de
Na + del medio externo hacia la hemolinfa, compensando la pérdida pasiva de iones inorgánicos (Pan et al., 2019). Las concentraciones séricas de iones se analizaron después de la interferencia de dsRNA Pv-miox. Los resultados mostraron que el estrés de baja salinidad provocó que los iones en el cuerpo se vieran obligados a perderse, pero el grupo de dsRNA Pvmiox ralentizó significativamente este fenómeno en comparación con el grupo de control. Además, los resultados de qRT-PCR mostraron que, después de la interferencia de dsRNA de Pv-miox, los niveles de expresión de los genes relacionados con el transportador de iones nka, nhe, ca y cam fueron significativamente más altos que los del grupo control. La posible razón es que Pv-miox está involucrado directa o indirectamente en el proceso de regulación osmótica bajo estrés de baja salinidad. La interferencia con dsRNA de Pv-miox conduce a un metabolismo debilitado de mioinositol, aumento de su síntesis y acumulación en los tejidos. Este fenómeno verificó una vez más que la síntesis de mioinositol y las vías metabólicas están involucradas en el proceso de regulación osmótica. Como efector osmótico, el mioinositol se acumula en las branquias bajo condiciones de
baja salinidad, promoviendo la actividad de las enzimas de transporte de iones y manteniendo así el equilibrio iónico en el cuerpo. También puede reducir la pérdida de iones inorgánicos causada por el estrés por baja salinidad y garantizar el transporte normal de iones (Bu et al., 2021, Cui et al., 2020, Nakanishi et al., 1989).
Adicionalmente, las branquias de P. vannamei son órganos clave para adaptarse a los cambios de salinidad. Cuando la salinidad externa cambia, P. vannamei depende no solo de poderosos mecanismos reguladores osmóticos sino también de otros mecanismos de adaptación fisiológica que desempeñan papeles clave en el proceso de adaptación a la salinidad (Esparza-Leal et al., 2019, Gao et al., 2016, Li et al., 2017). La vía de señalización Nrf2-Keap1 puede regular la transcripción de genes antioxidantes, incluyendo sod y gpx, para contrarrestar el estrés oxidativo causado por el estrés externo (Dawood et al., 2022, Ou y Liu, 2022, Sinha et al., 2015). En este estudio, la baja salinidad condujo a un aumento en la expresión de genes antioxidantes, lo que indica que el estrés oxidativo ocurrió en las branquias. Sin embargo, el nivel de expresión del gen antioxidante en el grupo Pv-miox dsRNA fue significativamente mayor que en el grupo control, lo que indica que la interferencia de Pv-miox dsRNA puede promover la resistencia al daño por estrés oxidativo a través de la vía de señalización nrf2 / sod / gpx El estrés oxidativo y el estrés del retículo endoplasmático están interrelacionados. Cuando el estrés oxidativo aumenta, el nivel de ROS en las células aumenta. Las ROS interfieren directa o indirectamente con la función del retículo endoplasmático y afectan el plegamiento y procesamiento de proteínas en el retículo endoplasmático (Zhang et al., 2019). El estrés del retículo endoplasmático implica la acumulación de proteínas mal plegadas o desplegadas, entre las que bip, ire1 y xbp1s son los factores distintivos del estrés del retículo endoplasmático (Bettigole y Glimcher, 2015, Oakes y Papa, 2015). En este estudio, el estrés por baja salinidad aumentó significativamente la expresión de los genes bip, ire1 y xbp1s, lo que indica que el estrés por baja salinidad causa estrés en el retículo endoplasmático a través de la vía de señalización bip / ire1 / xbp1s
Sin embargo, la interferencia del ARNdc de Pv-miox puede promover la vía de señalización bip / ire1 / xbp1s para regular el plegamiento de proteínas, reducir el número de proteínas mal plegadas y ayudar a las células a afrontar el estrés.
Los cambios de salinidad no solo causan estrés oxidativo y estrés del retículo endoplasmático, sino que también desencadenan la liberación de factores inflamatorios, activan células inmunes innatas y, en casos graves, incluso inician mecanismos de muerte celular para proteger a todo el organismo (El-Leithy et al., 2019, Gao et al., 2024, Schmitz et al., 2017). La relación entre la inflamación y la apoptosis es dinámica y compleja. NF-κB es un factor de transcripción que activa la citocina inflamatoria TNF-α (Wu y Zhou, 2010). JNK desempeña un papel en numerosas funciones fisiológicas, como la inflamación y la apoptosis (Xu et al., 2022). CytC desencadena una cascada
de genes de caspasa, lo que resulta en la apoptosis celular (Wang et al., 2020). En el presente estudio, los niveles aumentados de los genes nf- κ b, tnf- α y jnk indicaron que la baja salinidad indujo inflamación en las branquias de P. vannamei. La interferencia de ARNdc de Pv-miox aumentó la expresión de las vías nf- κ b, tnf-α y jnk relacionadas con la inflamación. La posible razón es que promueve la producción de factores inflamatorios, que pueden mantener la respuesta inmune mientras protegen a las células de la muerte prematura. Además, los niveles de expresión de los genes cytc, casp3 y casp-9 fueron significativamente menores después de la interferencia de ARNdc de Pvmiox que en el grupo control, verificando que la interferencia de ARNdc de Pv-miox aumentó la expresión de factores relacionados con la inflamación para proteger a las células de la muerte prematura. Bajo condiciones de estrés de baja salinidad, aunque la respuesta inflamatoria se activó, la respuesta apoptótica se inhibió efectivamente, lo que
Fig. 11. Diagrama mecanístico: efectos de la interferencia del ARNdc con Pv-miox sobre el estrés por baja salinidad en P. vannamei. El estrés agudo por baja salinidad afecta el equilibrio osmótico, lo que provoca estrés oxidativo, estrés del retículo endoplasmático, inflamación y apoptosis celular agravada. La interferencia del ARNdc con la vía metabólica del mioinositol puede aumentar la capacidad de regulación osmótica y la respuesta inmunitaria del camarón, ayudándolo así a adaptarse mejor a entornos de estrés por baja salinidad.
indica que el camarón blanco se adapta al estrés ambiental regulando el equilibrio entre las señales inflamatorias y apoptóticas. La interferencia de ARNdc de Pv-miox puede ayudar a los camarones a afrontar mejor el estrés por baja salinidad al potenciar la respuesta inflamatoria e inhibir la muerte celular prematura, manteniendo al mismo tiempo la homeostasis fisiológica. Este proceso puede implicar una activación moderada de la respuesta inmunitaria para evitar el daño celular causado por una respuesta inmunitaria excesiva. Por lo tanto, el estrés por baja salinidad puede alterar la homeostasis fisiológica de P. vannamei al inducir estrés oxidativo, estrés del retículo endoplasmático, inflamación y apoptosis. La interferencia de dsRNA puede aumentar la interacción entre estos factores relacionados con el sistema inmunológico y mejorar el estado adaptativo del camarón al estrés de baja salinidad.
Conclusión
Este estudio exploró el papel clave del metabolismo del mioinositol en la respuesta de P. vannamei al estrés por baja salinidad. La clonación e identificación del ADNc de longitud completa de Pv-miox reveló que el gen estaba altamente conservado a lo largo de la evolución y puede estar relacionado con la adaptación a la salinidad. Experimentos adicionales de interferencia de dsRNA revelaron que la acumulación de mioinositol en las branquias causada por la reducción del metabolismo de mioinositol era esencial para mantener el equilibrio de la presión osmótica celular. Además, el estrés oxidativo, el estrés del retículo endoplasmático y la respuesta inflamatoria inducida por el estrés por baja salinidad se regularon eficazmente en el grupo de interferencia de dsRNA de Pvmiox, previniendo la muerte celular excesiva.
En resumen, el metabolismo del mioinositol juega un papel importante en la adaptación de P. vannamei a ambientes de baja salinidad, especialmente en la regulación osmótica, el mantenimiento del equilibrio iónico y la respuesta inmune. El mecanismo específico se muestra en la figura (Fig. 11). El gen Pv-miox ayuda a los camarones a afrontar mejor el estrés ambiental en condiciones de baja salinidad al regular el metabolismo del mioinositol, lo que sugiere que este puede ser un objetivo potencial para mejorar la resistencia a la salinidad de los animales acuáticos•
Para más información sobre este artículo, contactar a: flhan@hainanu.edu.cn
Efectos de un co-cultivo inactivado por calor de Lactobacillus spp. (postbiótico) sobre el
rendimiento de crecimiento y la tolerancia al estrés ambiental en postlarvas de camarón blanco (Penaeus vannamei): síntesis de cuatro ensayos independientes
Autores:
Corneillie S.
Bancais M.
Tanguy C.
Cherel D.
sergecorneillie@gmail.com
Las fases larval y postlarval de la producción de camarón se caracterizan por una elevada sensibilidad a los factores de estrés ambiental, una supervivencia variable y una fuerte dependencia de la calidad fisiológica de las postlarvas (PL). Los aditivos funcionales se utilizan cada vez más durante estas etapas para mejorar el crecimiento, la robustez y la tolerancia al estrés. Entre ellos, los postbióticos (definidos como preparaciones de células microbianas inactivadas y/o sus metabolitos) han ganado gran interés debido a su estabilidad durante el procesamiento y almacenamiento del alimento, su composición predecible y su perfil de seguridad en comparación con los probióticos vivos.
Las preparaciones de Lactobacillus inactivadas por calor han demostrado mejorar el crecimiento, la competencia inmunitaria y la resistencia al estrés en diversas especies acuáticas, incluidos los camarones peneidos. Sin embargo, las respuestas suelen variar entre estudios, en parte debido a las diferencias en las condiciones experimentales y, de forma crítica, a la calidad intrínseca de las PL utilizadas al inicio de los ensayos. Las PL de
alta calidad normalmente presentan elevada supervivencia y potencial de crecimiento, dejando un margen limitado para detectar mejoras adicionales, especialmente bajo desafíos de estrés a corto plazo.
El presente artículo integra y analiza críticamente cuatro ensayos controlados realizados en Tailandia, dos en la Universidad de Kasetsart y dos en la Universidad Prince of Songkhla. Todos los ensayos evaluaron el mismo postbiótico de Lactobacillus inactivado por calor, pero presentaron diferencias marcadas en la calidad de las PL y en el desempeño basal.
El objetivo de esta síntesis es: (i) resumir los efectos del postbiótico sobre crecimiento y supervivencia; (ii) comparar los resultados de resistencia al estrés entre PL de baja y alta calidad, y (iii), discutir las implicaciones de la calidad de las PL para la interpretación de la eficacia de los postbióticos en los criaderos de camarón.
Materiales y métodos
Producto experimental
El postbiótico evaluado (Metalac®, STI
Biotechnology) consistió en una preparación doblemente fermentada e inactivada por calor de un co-cultivo de dos cepas de Lactobacillus: Lactobacillus rhamnosus y Lactobacillus farciminis
Las bacterias fueron inactivadas mediante tratamiento térmico, produciendo células no viables pero preservando componentes estructurales (por ejemplo, fracciones de pared celular) y metabolitos derivados de la fermentación. El producto se incorporó a los alimentos comerciales para larvas o postlarvas de camarón mediante recubrimiento superficial o recubrimiento post-extrusión, dependiendo del ensayo.
Sitios experimentales y calidad de las postlarvas
Se incluyeron cuatro ensayos independientes:
• Universidad de Kasetsart (SP1004 y SP1104): las PL se caracterizaron por bajo desempeño basal, crecimiento lento y baja supervivencia bajo condiciones control, lo que indica una calidad inicial subóptima.
• Universidad Prince of Songkhla (SP1001 y SP1103): las PL mostraron alta supervivencia basal y rápido crecimiento, reflejando alta calidad inicial.
Las PL se sembraron en etapas de desarrollo comparables (PL5) y se criaron hasta aproximadamente PL30 bajo condiciones estándar de criadero. En los ensayos de Kasetsart, las postlarvas se mantuvieron durante 20 días (PL5–PL30) en acuarios de vidrio de 80 L a una densidad de 25 camarones L-¹, equivalente a aproximadamente 2 000 postlarvas por tanque. Cada tratamiento dietético se evaluó con cinco réplicas y se probaron nueve tratamientos experimentales.
En los cuatro ensayos, los camarones recibieron una combinación de Artemia decapsulada y un alimento peletizado formulado con 42% de proteína cruda y 6% de lípidos crudos. La alimentación se realizó cuatro veces al día a intervalos regulares. La temperatura del agua se mantuvo a 29 °C y la salinidad a 25 ppt durante todo el período experimental.
En los ensayos de la Universidad Prince of Songkhla, los camarones se criaron bajo las mismas condiciones y esquema de alimentación, con la excepción de que se mantuvieron en acuarios de vidrio de 200 L a una densidad aproximada de 300 camarones m-².
Dietas y diseño experimental
Cada ensayo incluyó una dieta control sin suplementación y una o más dietas experimentales que contenían el postbiótico de Lactobacillus inactivado por calor en niveles de inclusión entre 0.5 y 5.0 kg t-¹ de alimento. La alimentación se realizó varias veces al día según la práctica local de los criaderos.
Rendimiento de crecimiento y supervivencia
El rendimiento de crecimiento se evaluó mediante mediciones del peso corporal individual y/o de la longitud al final del período de cría (generalmente PL30). La supervivencia se calculó como el porcentaje de PL restantes al momento de la cosecha en relación con el número inicial sembrado.
Pruebas de resistencia al estrés
Al final del período de alimentación, subgrupos de PL se sometieron a desafíos agudos de estrés, según el ensayo:
• Estrés de salinidad: exposición a salinidad baja o alta (por ejemplo, 0 o 40 ppt).
• Estrés térmico: exposición a temperatura baja o alta (por ejemplo, 15 o 35 °C).
• Estrés químico: exposición a formalina en concentraciones graduales.
La supervivencia se registró después de un período fijo de exposición (generalmente 24 h). En ensayos seleccionados se analizaron indicadores bioquímicos o moleculares de estrés, como la actividad de glutatión peroxidasa (GPx) o la expresión de la proteína de choque térmico 70 (HSP70).
Análisis estadístico
Dentro de cada ensayo, los datos se analizaron mediante análisis de varianza (ANOVA), seguido de comparaciones post hoc cuando correspondía. Las diferencias se consideraron significativas con p < 0.05, según se reportó en los informes originales.
Resultados
Desempeño basal y calidad de las PL
Se observaron diferencias marcadas en el desempeño basal entre las instituciones. En los ensayos de Kasetsart, la supervivencia del control a PL30 osciló entre aproximadamente 46–62%, con ganancias de peso absolutas muy bajas. En contraste, los grupos control de Songkhla mostraron tasas de supervivencia cercanas al 90% o superiores y pesos finales sustancialmente mayores.
Rendimiento de crecimiento bajo condiciones normales
En los cuatro ensayos, la suplementación dietética con el postbiótico de Lactobacillus inactivado por calor mejoró el rendimiento de crecimiento respecto al control. Las respuestas más consistentes y reproducibles se observaron con un nivel de inclusión de
aproximadamente 1 kg t-¹ de alimento.
En los ensayos de Kasetsart, las mejoras de crecimiento fueron moderadas en términos absolutos, pero biológicamente relevantes dada la baja línea base. En los ensayos de Songkhla, las mejoras relativas en ganancia de peso y eficiencia alimenticia fueron frecuentemente pronunciadas, incluyendo reducciones sustanciales en el factor de conversión alimenticia.
Supervivencia bajo condiciones normales de cría
Las respuestas de supervivencia dependieron fuertemente de la calidad de las PL. En PL de baja calidad en Kasetsart, la suplementación con postbiótico mejoró significativamente la supervivencia, en algunos casos incrementándola de menos del 50% en el control a más del 80% en los niveles óptimos de inclusión. En PL de alta calidad en Songkhla, la supervivencia ya era elevada en los grupos control y generalmente se mantuvo, con diferencias menores o inconsistentes entre tratamientos.
Resistencia al estrés
Los resultados de los desafíos de estrés mostraron la interacción más clara con la calidad de las PL. En los ensayos de Kasetsart, la suplementación con postbiótico mejoró la supervivencia bajo estrés de salinidad y temperatura, confirmando una mayor robustez de las PL de baja calidad.
En los ensayos de Songkhla, la supervivencia bajo condiciones de estrés ya era muy alta en los grupos control y los beneficios adicionales del postbiótico fueron limitados o no estadísticamente significativos.
Tabla 1. Resumen de los cuatro ensayos de alimentación que evaluaron un postbiótico de Lactobacillus inactivado por calor en postlarvas de Penaeus vannamei
de Kasetsart
SP1004
SP1104
PL10–PL30 0, 0.5–5.0 kg t-¹ Crecimiento, supervivencia, estrés salinidad y temperatura
de Kasetsart Baja PL10–PL30 0, 0.5–5.0 kg t-¹ Crecimiento, supervivencia, estrés salinidad y temperatura
SP1001 Universidad Prince of Songkhla Alta PL10–PL30 0, 0.5–5.0 kg t-¹ Crecimiento, supervivencia, estrés por formalina y salinidad
SP1103
Universidad Prince of Songkhla Alta PL10–PL30 0, 0.5–1.0 kg t-¹ Crecimiento, FCR, estrés por formalina y salinidad
PRODUCCIÓN
Los análisis de biomarcadores no demostraron de forma consistente la activación de las vías clásicas de estrés (por ejemplo, HSP70), lo que sugiere que los beneficios observados fueron mediados por mecanismos fisiológicos más amplios.
Discusión
El análisis integrado de estos cuatro ensayos demuestra claramente que la eficacia de un postbiótico de Lactobacillus inactivado por calor en postlarvas de camarón está fuertemente modulada por la calidad inicial de las PL. Si bien los beneficios en el rendimiento de crecimiento se observaron de manera consistente en todos los estudios, la detectabilidad y la magnitud de los efectos sobre la supervivencia y la resistencia al estrés dependieron de la robustez basal de las PL.
En poblaciones de PL de baja calidad, como las utilizadas en los ensayos de la Universidad de Kasetsart, la supervivencia y el crecimiento basales fueron pobres, lo que dejó un amplio margen para intervenciones funcionales. En estas condiciones, la suplementación con postbióticos se tradujo en mejoras biológicamente y económicamente significativas tanto en la supervivencia como en la tolerancia al estrés. Estos resultados son particularmente relevantes para los criaderos comerciales que enfrentan una calidad variable de PL, presión de enfermedades o inestabilidad ambiental estacional (por ejemplo, fluctuaciones de salinidad y temperatura durante la temporada de lluvias).
Por el contrario, las PL de alta calidad utilizadas en los ensayos de la Universidad Prince of Songkhla ya presentaban alta supervivencia y fuerte crecimiento bajo condiciones control. En estos casos, la suplementación con postbióticos mejoró principalmente la eficiencia de crecimiento, mientras que los ensayos de estrés frecuentemente alcanzaron un efecto techo, ocultando beneficios protectores adicionales. Esta observación no invalida la eficacia del postbiótico, sino que pone de relieve las limitaciones de las pruebas de estrés agudo cuando se aplican a poblaciones de PL robustas.
Los presentes hallazgos son coherentes
Tabla 2. Resumen de las respuestas de crecimiento bajo condiciones normales de cría
Final
Tabla 3. Resumen de las respuestas de supervivencia bajo condiciones normales de cría
4. Resultados de los desafíos de estrés en relación con la calidad de PL
C)
(15 degr. C)
con informes previos que describen mejoras en el crecimiento, la competencia inmunitaria y la resistencia al estrés en camarones alimentados con preparaciones de Lactobacillus inactivadas o muertas por calor. Estudios en Marsupenaeus japonicus y Penaeus vannamei han demostrado un aumento de la supervivencia bajo estrés por salinidad y manipulación, así como la modulación de parámetros inmunitarios no específicos, particularmente bajo condiciones de cría subóptimas (Tung et al., 2009; Wu et al., 2024; Goh et al., 2022).
En conjunto, estos datos refuerzan el concepto de que los postbióticos funcionan principalmente como potenciadores de la resiliencia, con los beneficios más marcados cuando los animales están expuestos a
desafíos ambientales o fisiológicos.
Posicionamiento de los postbióticos frente a los probióticos
Los resultados de estos ensayos respaldan el posicionamiento estratégico de los productos de Lactobacillus inactivados por calor como postbióticos, en lugar de probióticos clásicos. A diferencia de los probióticos vivos, los postbióticos no dependen de la colonización intestinal ni de la actividad metabólica in situ. En cambio, sus efectos funcionales están mediados por patrones moleculares asociados a microbios (MAMPs), componentes de la pared celular y metabolitos derivados de la fermentación que interactúan con las vías inmunitarias y metabólicas del hospedador.
Tabla
(Kasetsart vs Prince)
Esta distinción es especialmente relevante en sistemas de criadero de camarón, donde las condiciones de procesamiento del alimento, las prácticas de desinfección del agua y el tránsito intestinal rápido pueden limitar la viabilidad y la consistencia de los probióticos vivos. Los postbióticos ofrecen una estabilidad superior, una dosificación predecible y compatibilidad con procesos de peletizado/extrusión y almacenamiento a largo plazo, al tiempo que proporcionan mejoras medibles en el rendimiento de crecimiento y la robustez.
Implicaciones para la práctica en criaderos Estos cuatro ensayos subrayan la necesidad de caracterizar explícitamente la calidad de las PL al inicio de la cría o del diseño experimental. Sin esta estratificación, las comparaciones entre estudios o resultados productivos pueden conducir a conclusiones
erróneas sobre la eficacia de los aditivos. Incluso puede plantearse la evaluación de la calidad de las PL antes de la siembra en cualquier estanque para predecir los resultados productivos.
Para futuras evaluaciones de postbióticos en criaderos de camarón, se recomienda el uso de variables complementarias (como marcadores inmunitarios, histología intestinal, desafíos patógenos o análisis de tiempo hasta la mortalidad), especialmente cuando se trabaja con PL de alta calidad.
En términos prácticos, los postbióticos de Lactobacillus inactivados por calor representan una herramienta robusta y escalable para mejorar la eficiencia de crecimiento y amortiguar las pérdidas de rendimiento asociadas a la variabilidad en la calidad de las PL y al estrés ambiental•
Referencias
Tung, H.T. et al. (2009). Effects of heat killed Lactobacillus plantarum on growth performance, stress resistance and immune response of kuruma shrimp (Marsupenaeus japonicus). Aquaculture Science, 57, 175–184.
Wu, T. et al. (2024). Effect of dietary supplement of inactivated Lactobacillus plantarum on growth performance, immune response, disease resistance, and intestinal microbiota in Penaeus vannamei. Journal of Oceanology and Limnology, 42, 676–694.
Kasetsart University. SP1004 and SP1104 PL shrimp trial reports (2011).
Prince of Songkhla University. SP1001 and SP1103 PL shrimp trial reports (2011).
Goh, J.X.H. et al. (2022). Probiotics, paraprobiotics and postbiotics in aquaculture: mechanisms and applications. Reviews in Aquaculture.
Técnicas recomendadas para la biorremediación del estuario del Guayas y su área de influencia el perfil costero–marino de la provincia de Santa Elena
Autor:
Leonardo S. Maridueña - Dirección de Ambiente de la Cámara Nacional de Acuacultura lmariduena@cna-ecuador.com
El estuario del río Guayas constituye uno de los ecosistemas estuarinos más importantes del Ecuador debido a su elevada productividad biológica y su relevancia ambiental, social y económica. Sin embargo, este sistema se encuentra sometido a una intensa presión antrópica derivada de la descarga de aguas residuales domésticas, efluentes industriales y mineros, así como de la escorrentía superficial proveniente de actividades agrícolas que transporta pesticidas y plaguicidas hacia el cuerpo estuarino. Estas fuentes de contaminación han provocado un deterioro significativo de la calidad del agua y los sedimentos, evidenciado por elevados valores de demanda bioquímica de oxígeno (DBO), disminución del oxígeno disuelto, alteraciones del pH, acumulación de metales pesados como cadmio (Cd), plomo (Pb) y mercurio (Hg); presencia de aceites y grasas, y una elevada carga microbiológica caracterizada por bacterias patógenas, incluidos coliformes fecales.
El presente artículo analiza la aplicación de técnicas de biorremediación como una alternativa ambientalmente sostenible para la recuperación del estuario del río Guayas y la mitigación de impactos hacia el Golfo de Guayaquil y el perfil costero–marino de la provincia de Santa Elena. Se evalúan estrategias como la bioestimulación y bioacumulación microbiana para la reducción de la DBO y el control de bacterias patógenas, la biosorción microbiana y algal para la inmovilización de metales pesados asociados a descargas mineras, y la fitorremediación en zonas intermareales
para la estabilización de sedimentos contaminados por metales y agroquímicos.
La aplicación integrada de estas técnicas se presenta como una opción viable y de bajo impacto ambiental para el cumplimiento del Acuerdo Ministerial 097-A del Ministerio del Ambiente del Ecuador, específicamente de lo establecido en sus Tablas 2 y 10.
Los estuarios son ecosistemas altamente dinámicos que actúan como zonas de
transición entre ambientes continentales y marinos, caracterizados por una elevada productividad biológica y una compleja interacción de procesos físicos, químicos y biológicos. No obstante, su ubicación estratégica los convierte en receptores naturales de contaminantes provenientes de actividades urbanas, industriales, mineras y agrícolas.
El estuario del río Guayas es uno de los ecosistemas más grandes de la costa oeste de Sudamérica y representa uno de los sistemas estuarinos más relevantes del litoral ecuatoriano. La degradación progresiva de su calidad ambiental no solo afecta al propio estuario, sino que genera impactos acumulativos hacia el Golfo de Guayaquil y el perfil costero–marino de la provincia de Santa Elena, evidenciando la necesidad de enfoques integrados de gestión ambiental y recuperación ecológica.
Dinámica hidrológica del estuario del Guayas
La dinámica hidrológica está controlada por la interacción entre el caudal fluvial, las mareas semidiurnas del Golfo de Guayaquil, las
corrientes marinas y la variabilidad estacional de las precipitaciones. Estos procesos determinan el transporte, la dispersión y la acumulación de contaminantes, así como la distribución del oxígeno disuelto y el pH.
Durante la época lluviosa, el incremento del caudal fluvial intensifica el aporte de sedimentos y contaminantes, mientras que eventos climáticos como El Niño exacerban la estratificación de la columna de agua y la disminución del oxígeno disuelto. La comprensión de esta dinámica es fundamental para la planificación y efectividad de las estrategias de biorremediación.
Perfil costero–marino de la provincia de Santa Elena
Este perfil costero–marino se caracteriza por una mayor energía hidrodinámica, influenciada por corrientes oceánicas, oleaje y procesos de mezcla marina. A través del Golfo de Guayaquil, este perfil se encuentra funcionalmente conectado al estuario del río Guayas, lo que lo hace vulnerable a impactos indirectos derivados del transporte de contaminantes estuarinos. La calidad ambiental de esta zona es crítica para actividades como la pesca, la acuicultura y el turismo.
Marco normativo ambiental
La regulación de la calidad ambiental del agua superficial y de las descargas de efluentes en el Ecuador se establece mediante el Acuerdo Ministerial 097-A del Ministerio del Ambiente, el cual define los límites máximos permisibles para parámetros fisicoquímicos, microbiológicos y tóxicos.
En particular, la Tabla 2 regula la calidad del agua superficial, mientras que la Tabla 10 establece los límites para descargas de efluentes al recurso hídrico. Entre los parámetros regulados de mayor relevancia para el estuario del Guayas se incluyen la demanda bioquímica de oxígeno (DBO5), oxígeno disuelto, pH, coliformes fecales, metales pesados (Cd, Pb y Hg), aceites y grasas e hidrocarburos totales de petróleo. La superación de estos límites en diversos sectores del estuario evidencia un deterioro ambiental que justifica la implementación de estrategias de remediación.
Problemática ambiental del estuario
El estuario del Guayas enfrenta una problemática ambiental compleja y multifactorial como resultado del crecimiento urbano acelerado, la expansión de actividades industriales y mineras, y el desarrollo intensivo de la agricultura en su cuenca hidrográfica. Estas presiones antrópicas han alterado de manera significativa la calidad del agua y los sedimentos, comprometiendo las funciones ecológicas del estuario y los servicios ecosistémicos que brinda a las poblaciones humanas y se las define a continuación:
Contaminación orgánica y demanda bioquímica de oxígeno (DBO)
La descarga continua de aguas residuales domésticas, en muchos casos sin tratamiento o con tratamiento insuficiente, constituye una de las principales fuentes de contaminación orgánica en el estuario del Guayas. Estas descargas incrementan de forma significativa los valores de demanda bioquímica de oxígeno (DBO), superando los límites máximos permisibles establecidos en la normativa ambiental ecuatoriana para cuerpos de agua superficiales.
Los altos valores de DBO generan una elevada demanda de oxígeno disuelto, provocando condiciones de hipoxia o anoxia, especialmente en zonas de baja renovación hidráulica y en sedimentos finos. Estas condiciones afectan negativamente a organismos acuáticos sensibles, reducen la biodiversidad y alteran los procesos biogeoquímicos naturales del sistema estuarino.
Tabla 2, del Acuerdo Ministerial 097A
Contaminación microbiológica y presencia de bacterias patógenas
La contaminación microbiológica del estuario del Guayas se manifiesta por la elevada presencia de bacterias patógenas, particularmente coliformes fecales, los cuales son indicadores de contaminación de origen fecal. Esta problemática está directamente relacionada con la descarga de aguas residuales domésticas y urbanas sin un tratamiento adecuado.
La presencia de coliformes fecales y otros microorganismos patógenos representa un riesgo significativo para la salud pública, especialmente para comunidades que dependen del estuario para actividades como la pesca artesanal, la acuicultura y el uso recreativo del agua. Asimismo, la normativa ambiental ecuatoriana establece límites específicos para coliformes fecales, los cuales son frecuentemente excedidos en sectores del estuario, evidenciando una degradación de la calidad sanitaria del recurso hídrico.
Metales pesados y descargas de origen minero e industrial
Las actividades mineras, así como ciertos procesos industriales desarrollados en la cuenca del Guayas, aportan metales pesados como cadmio (Cd), plomo (Pb) y mercurio (Hg) al sistema estuarino. Estos contaminantes se caracterizan por su persistencia, toxicidad y capacidad de bioacumulación y biomagnificación a lo largo de la cadena trófica.
Parámetros de calidad del agua superficial para preservación de la vida acuática
(Extracto técnico – Tabla 2, AM 097-A)
Parámetro
Demanda bioquímica de oxígeno (DBO5)
Oxígeno disuelto (OD)
Coliformes fecales
Cadmio (Cd)
Plomo (Pb)
Mercurio total (Hg)
Hidrocarburos totales de petróleo (HTP)
Aceites y grasas
Sólidos suspendidos totales
En el estuario, los metales pesados tienden a asociarse con partículas finas y materia orgánica, acumulándose principalmente en los sedimentos. Cambios en las condiciones fisicoquímicas, como variaciones de pH o potencial redox, pueden favorecer su remobilización, incrementando su biodisponibilidad y riesgo ecológico. La normativa ambiental ecuatoriana establece límites máximos permisibles para estos metales, los cuales pueden ser superados en zonas influenciadas por descargas mineras y efluentes industriales.
Contaminación por aceites, grasas e hidrocarburos
La presencia de aceites, grasas e hidrocarburos en el estuario del Guayas se asocia principalmente a actividades portuarias, industriales y al transporte fluvial. Estos contaminantes forman películas superficiales que reducen el intercambio gaseoso entre el agua y la atmósfera, afectando la oxigenación del sistema.
Además, muchos hidrocarburos presentan propiedades tóxicas y persistentes, acumulándose en los sedimentos y afectando a organismos bentónicos. La normativa ecuatoriana regula la descarga de hidrocarburos totales; sin embargo, su presencia recurrente evidencia deficiencias en el control de efluentes.
Pesticidas, plaguicidas y escorrentía agrícola
La escorrentía superficial proveniente de áreas agrícolas de la cuenca del río Guayas transporta pesticidas y plaguicidas hacia el estuario, tanto disueltos en el agua como asociados a sedimentos. Estos compuestos pueden generar efectos tóxicos agudos y crónicos sobre organismos acuáticos, alterar procesos reproductivos y reducir la diversidad biológica.
La persistencia de ciertos agroquímicos y su interacción con otros contaminantes presentes en el estuario pueden potenciar efectos sinérgicos, incrementando el riesgo ecológico. Este tipo de contaminación difusa es de difícil control y representa uno de los mayores desafíos para la gestión ambiental del estuario.
Contaminación de sedimentos y efectos acumulativos
Tabla 10, del Acuerdo Ministerial 097A
Límites máximos permisibles para descargas de efluentes al recurso agua marina.
(Extracto técnico – Tabla 10, AM 097-A)
Parámetro
Unidad Límite máximo permisible
Demanda bioquímica de oxígeno (DBO5) mg/L ≤ 200
Demanda química de oxígeno (DQO) mg/L ≤ 500
Oxígeno disuelto (OD) mg/L No aplica
pH
Coliformes fecales
Aceites y grasas
Cadmio (Cd)
Plomo (Pb)
Mercurio total (Hg)
Hidrocarburos totales de petróleo (HTP)
Sólidos suspendidos totales
Los sedimentos del estuario del Guayas actúan como reservorios de contaminantes orgánicos, metales pesados y agroquímicos. La acumulación prolongada de estos compuestos genera efectos acumulativos que afectan la calidad del hábitat bentónico y la salud del ecosistema en su conjunto.
Procesos naturales como la resuspensión de sedimentos durante eventos de marea, lluvias intensas o actividades de dragado pueden liberar contaminantes nuevamente a la columna de agua, prolongando los impactos ambientales y dificultando los procesos de recuperación natural del estuario.
Técnicas de biorremediación aplicables al estuario del Guayas
La biorremediación se presenta como una alternativa ambientalmente sostenible frente a métodos convencionales de remediación. La bioestimulación microbiana, mediante la optimización de nutrientes, oxígeno y pH, permite acelerar la degradación de materia orgánica, reducir la demanda bioquímica de oxígeno (DBO) e incrementar los niveles de oxígeno disuelto. La bioaumentación, mediante la inoculación de consorcios microbianos especializados, mejora la degradación de compuestos orgánicos persistentes y contribuye al control de bacterias patógenas por competencia biológica.
La biosorción microbiana y algal constituye una herramienta eficaz para la inmovilización de metales pesados asociados a descargas mineras, reduciendo su biodisponibilidad y toxicidad. Por su parte, la fitorremediación,
30
aplicada en zonas intermareales mediante especies halófitas, permite la estabilización de sedimentos contaminados y la reducción de la movilidad de metales y agroquímicos. Estas técnicas pueden complementarse con bioturbación controlada y enmiendas orgánicas para mejorar las condiciones redox del sedimento.
La complejidad del estuario del Guayas requiere la aplicación de estrategias integradas de biorremediación, adaptadas a las condiciones de salinidad variable, dinámica mareal y diversidad de contaminantes presentes en agua y sedimentos, que pueden ser:
Bioestimulación microbiana
La bioestimulación consiste en la adición controlada de nutrientes esenciales (nitrógeno y fósforo), junto con la optimización de condiciones ambientales como oxígeno disuelto y pH, con el fin de estimular la actividad de microorganismos autóctonos degradadores.
En el estuario del Guayas, esta técnica resulta particularmente efectiva para:
• Reducir la DBO asociada a materia orgánica biodegradable
• Acelerar la degradación de aceites, grasas e hidrocarburos
• Disminuir la persistencia de pesticidas orgánicos
Además, el incremento de la actividad microbiana benéfica puede generar competencia biológica, contribuyendo a la reducción de bacterias patógenas, incluidos coliformes fecales.
Bioaumentación microbiana
La bioaumentación implica la inoculación de consorcios microbianos especializados, capaces de tolerar altas cargas contaminantes y condiciones estuarinas variables. Esta técnica es especialmente útil en zonas donde la microbiota nativa ha sido afectada por contaminación crónica.
Su aplicación permite:
• Mejorar la degradación de compuestos orgánicos persistentes
• Aumentar la eficiencia en la reducción de DBO
• Complementar el control microbiológico del sistema
• Biosorción microbiana y algal de metales pesados
La biosorción (proceso fisicoquímico natural donde biomasas vivas o muertas, como algas, hongos o bacterias, capturan y concentran contaminantes) permitiría inmovilizar metales pesados mediante mecanismos de adsorción en la pared celular y exopolisacáridos. En el estuario del Guayas, esta técnica es especialmente relevante para mitigar impactos de descargas mineras que aportan Cd, Pb y Hg.
La biosorción reduce la biodisponibilidad de los metales, minimizando su toxicidad y el riesgo de bioacumulación.
Fitorremediación en zonas intermareales
La fitorremediación emplea vegetación halófita adaptadas a concentraciones salinas ya sea para condiciones estuarinas o marinas, como por ejemplo manglar y algas. Las macroalgas pueden absorber contaminantes, funcionando como agentes de biorremediación naturales al limpiar aguas residuales y mejorar ecosistemas costeros degradados. A través de procesos de biosorción, absorben nutrientes como nitrógeno y fósforo, metales pesados, compuestos orgánicos e incluso retienen una porción significativa del CO2 oceánico. Para la fitorremediación puede usarse también la vegetación que crece en el borde costero y su función es para:
• Estabilizar sedimentos contaminados
• Extraer o inmovilizar metales pesados
• Reducir la movilidad de pesticidas y plaguicidas
• Esta técnica resulta adecuada para
márgenes y zonas intermareales del estuario o zona costera aportando beneficios adicionales como la recuperación del hábitat y la protección contra erosión.
Técnicas complementarias
La bioturbación controlada mediante organismos bentónicos y el uso de enmiendas orgánicas biochar, que es un material sólido, poroso y rico en carbono, obtenido al calentar residuos orgánicos (biomasa) a altas temperaturas en ausencia de oxígeno (proceso conocido como pirólisis) y compost estabilizado, mejoran las condiciones redox del sedimento, favorecen la actividad microbiana beneficiosa e incrementan la remoción de contaminantes.
Discusión comparativa: estuario del Guayas y costa de Santa Elena
Esquema de conectividad hidrológica entre el Golfo de Guayaquil y el perfil costero–marino de la provincia de Santa Elena, evidenciando la dispersión de contaminantes mediante mareas, corrientes y transporte de sedimentos.
Estructura conceptual:
Estuario del Guayas
↓ (mareas y corrientes)
Golfo de Guayaquil
↓
Perfil costero de Santa Elena
(impacto en pesca, acuicultura, turismo) Discusión comparativa:
Estuario del Guayas vs. perfil costero–marino de Santa Elena
La relación entre el estuario del Guayas y el perfil costero–marino de la provincia de Santa Elena debe analizarse desde una perspectiva sistémica, considerando que ambos ambientes se encuentran funcionalmente conectados a través del Golfo de Guayaquil. Sin embargo, presentan diferencias significativas en términos de dinámica hidrológica, niveles de contaminación y capacidad de asimilación ambiental.
El estuario del Guayas actúa como un sistema receptor primario de contaminantes, caracterizado por elevadas cargas de materia orgánica, bacterias patógenas, metales pesados, aceites y compuestos agroquímicos. Su dinámica estuarina, con zonas de baja energía hidrodinámica y altos
tiempos de residencia del agua, favorece la acumulación de contaminantes en los sedimentos y la ocurrencia de condiciones hipóxicas, especialmente cuando se superan los límites establecidos en el Acuerdo Ministerial 097-A.
En contraste, el perfil costero–marino de la provincia de Santa Elena presenta una mayor energía hidrodinámica, influenciada por corrientes marinas, oleaje y procesos de mezcla oceánica, lo que permite una mayor dispersión y dilución de contaminantes. No obstante, esta capacidad de dispersión no elimina el riesgo ambiental, ya que los contaminantes transportados desde el estuario pueden afectar ecosistemas marinos, zonas de pesca y áreas turísticas, especialmente en períodos de alta descarga fluvial o durante eventos climáticos extremos como El Niño.
La elevada productividad primaria del Golfo de Guayaquil, dominada por fitoplancton y microalgas bentónicas, representa un alto potencial ecológico para la implementación de procesos de biorremediación. Estas comunidades algales pueden ser utilizadas como agentes de biosorción de nutrientes y metales, reforzando estrategias de remediación basadas en organismos autóctonos.
Los ecosistemas de manglar del Golfo de Guayaquil, ampliamente descritos por el CAAM (1996), funcionan como sistemas naturales de biorremediación de alta eficiencia ecológica. Sus sedimentos anóxicos favorecen procesos de desnitrificación, mientras que la rizosfera del manglar promueve la inmovilización de metales pesados y la degradación microbiana de compuestos orgánicos. Desde el punto de vista de la biorremediación, el estuario del Guayas constituye un sitio prioritario de intervención, donde las técnicas biológicas pueden aplicarse de manera más controlada y eficiente. En cambio, en el perfil costero de Santa Elena, las estrategias deben enfocarse principalmente en la prevención, el control de fuentes contaminantes y el monitoreo oceanográfico, debido a la mayor complejidad y escala del ambiente marino.
Esta comparación evidencia que la recuperación ambiental del estuario del
Guayas no solo es un objetivo local, sino una condición necesaria para la protección ambiental del Golfo de Guayaquil y del perfil costero de Santa Elena, reforzando la necesidad de un enfoque integrado estuario–perfil costero.
Conclusiones
La biorremediación representa una alternativa técnica viable, sostenible y alineada con la normativa ambiental ecuatoriana para la recuperación del estuario del río Guayas. Su aplicación integrada permite mejorar la calidad del agua, reducir riesgos sanitarios, disminuir la biodisponibilidad de metales pesados y mitigar impactos hacia el Golfo de Guayaquil y la costa de Santa Elena, contribuyendo a la conservación de los ecosistemas estuarinos y costeros del Ecuador•
Bibliografía consultada
APHA. (2017). Standard Methods for the Examination of Water and Wastewater. CAAM (1996). Sistemas biofísicos en el Golfo de Guayaquil.
Cifuentes, L. A., Coffin, R. B., & Solórzano, L. (1996). Isotopic and elemental variations of carbon and nitrogen in a mangrove estuary. Estuarine, Coastal and Shelf Science, 43(6), 781–800.
Gadd, G. M. (2010). Metals, minerals and microbes: geomicrobiology and bioremediation. Microbiology, 156(3), 609–643. https://doi.org/10.1006/ecss.1996.0099
INOCAR. (2018). Caracterización oceanográfica del Golfo de Guayaquil. Instituto Oceanográfico de la Armada, Ecuador.
Ministerio del Ambiente del Ecuador. (2015). Acuerdo Ministerial 097-A: Norma de calidad ambiental y de descarga de efluentes al recurso agua.
Muñoz, E., & Rivera, J. (2010). Dinámica sedimentaria del Golfo de Guayaquil y su influencia en ecosistemas costeros. Revista Geológica de Chile, 37(2), 325–340.
Perelo, L. W. (2010). In situ and bioremediation of organic pollutants in aquatic sediments. Journal of Hazardous Materials, 177(1–3), 81–89.
Stevenson, M. R. (1981). Seasonal variations of the Gulf of Guayaquil, Ecuador. Journal of Geophysical Research, 86(C6), 5073–5080. https://doi.org/10.1029/JC086iC
Stevenson, M. R. (1981). Seasonal variations of the Gulf of Guayaquil. Journal of Geophysical Research.
Twilley, R. R. et al. (2001). The Gulf of Guayaquil and the Guayas River estuary. Springer.
Twilley, R. R., Cárdenas, W., Rivera-Monroy, V. H., & Espinoza, J. (2001). The Gulf of Guayaquil and the Guayas River estuary, Ecuador. In Coastal marine ecosystems of Latin America (pp. 245–263). Ed. Springer. Vidali, M. (2001). Bioremediation: An overview. Pure and Applied Chemistry, 73(7), 1163–1172.
75 71 79
ESTADÍSTICAS ÍNDICE
Edición 169 - Febrero 2026
Exportaciones de camarón
Reporte de mercado de China
Reporte de mercado de EE. UU.
CAMARÓN Estadísticas de Comercio Exterior
Elaborado por: María Andrea Dicindio - Subgerente de Comercio Exterior de la Cámara Nacional de Acuacultura
EVOLUCIÓN DE LAS EXPORTACIONES: 2010 AL 2025
Las exportaciones de camarón muestran una tendencia creciente sostenida entre 2010 y 2025, tanto en volumen como en valor. El volumen exportado aumentó de 151 mil TM en 2010 a cerca de 1,4 millones de TM en 2025, evidenciando una fuerte expansión productiva. A partir de 2017 el crecimiento se acelera y se consolida desde 2021. En términos de valor, las exportaciones pasaron de USD 850 millones a aproximadamente USD 7.474 millones. En 2022 se registra un pico importante, superando los USD 7.000 millones, impulsado por mejores precios internacionales. Pese a leves ajustes en 2023 y 2024, el sector mantiene una trayectoria sólida y estratégica para la economía.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
ESTADÍSTICAS
MILLONES DE LIBRAS EXPORTADAS: COMPARATIVO MENSUAL 2021 AL 2025
Las exportaciones de camarón han mostrado un incremento constante desde 2021 hasta 2025 en casi todos los meses, con algunos meses alcanzando picos considerables.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
El gráfico muestra un crecimiento progresivo de las exportaciones mensuales de camarón entre 2021 y 2025. En todos los meses, 2025 registra los mayores volúmenes, consolidando una tendencia positiva frente a años anteriores. Se observan picos estacionales claros en mayo, junio y noviembre, meses que concentran los mayores embarques. Durante 2023 y 2024 los volúmenes se mantienen altos, aunque con ligeras variaciones mensuales. El año 2021 presenta los niveles más bajos del período, evidenciando la expansión sostenida del sector en los años posteriores. En conjunto, el comportamiento mensual confi rma una mayor estabilidad y capacidad exportadora hacia 2025.
PARTICIPACIÓN POR MERCADOS DESTINO: DICIEMBRE 2024 VS DICIEMBRE 2025
China se mantiene como el principal destino de las exportaciones durante el mes de diciembre 2025 vs el mismo mes en 2024, incrementando su participación de 52,6 % en 2024 a 56,1 % en 2025. Estados Unidos y Europa registran una reducción en su participación, pasando de 21,8 % a 18,8 % y de 18,2 % a 15,8 %, respectivamente.
El resto de Asia muestra un crecimiento moderado, aumentando de 4,6 % a 5,7 %, reflejando una mayor diversificación regional. América también incrementa levemente su participación, de 1,9 % a 2,6 %. África y Oceanía mantienen una participación marginal y estable respecto al año anterior.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
PRINCIPALES PAÍSES DESTINOS (MILLONES DE LIBRAS):
ENERO - DICIEMBRE (2024 VS 2025)
La tendencia general muestra crecimiento en todos los países/regiones, destacando principalmente a Francia y España, presentando un crecimiento del 50% y 35% en el volumen exportado respectivamente. Por otro lado, se observa a Estados Unidos con un aumento del 26%, pasando de 473 a 597 millones de libras entre el período enero - diciembre del 2025 versus mismo período del 2024.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
EVOLUCIÓN DE EXPORTACIONES (MILLONES DE LIBRAS):
CHINA Y EE. UU. ENERO - DICIEMBRE 2025
El gráfico muestra que China mantiene volúmenes significativamente mayores que Estados Unidos durante todos los meses del año. Las exportaciones a China alcanzan su máximo en mayo (181) y presentan otro pico relevante en noviembre (149), con una caída marcada entre julio y septiembre. Estados Unidos muestra volúmenes más bajos pero relativamente estables, con su mayor nivel en octubre (61) y un mínimo en julio (38). Ambos mercados evidencian una estacionalidad similar, con descensos a mitad de año y recuperación hacia el último trimestre.
Fuente: Estadistic S.A.
Elaborado por: Cámara Nacional de Acuacultura
Importación de camarón de China
Autor: Erwin Termaat Kontali Shrimp
erwin.termaat@kontali.com www.kontali.com
El volumen de importaciones de camarones de China se fortaleció aún más en diciembre, alcanzando las 93,200 toneladas, lo que supone un aumento del 8% con respecto al mes anterior, pero un descenso del 8% con respecto al año anterior. El valor de las importaciones aumentó hasta los 528,8 millones de dólares, lo que supone un aumento del 7% con respecto al mes anterior, pero un descenso del 6% con respecto al año anterior, lo que refleja un mercado que se mantuvo activo, pero muy sensible a los precios. El precio medio de importación bajó ligeramente hasta los 5,67 dólares/kg (una caída de 0,09 dólares/ kg respecto al mes anterior), aunque se mantuvo 0,13 dólares/kg por encima del año anterior, lo que pone de relieve que el valor de las importaciones siguió estando más respaldado por los elevados costes de sustitución que por el crecimiento del volumen.
En el conjunto del año, China importó 988,740 toneladas en 2025 (una caída de aproximadamente un 1% respecto al año anterior). Sin embargo, el precio medio de importación aumentó hasta los 5,38 dólares/kg (desde los 5,07 dólares/ kg en 2024), lo que elevó el valor total de las importaciones a 5,320 millones de dólares (un aumento de alrededor del 5% interanual). Esta combinación apunta a una demanda subyacente ampliamente estable, un comportamiento de compra disciplinado y un crecimiento del valor impulsado por los costes, más que a una expansión impulsada por la demanda.
Rendimiento por especies
El suministro de camarones Vannamei alcanzó las 74,250 toneladas equivalentes a camarones vivos (LSE) en diciembre, lo
que supone un aumento del 5% respecto al mes anterior y del 3% respecto al año anterior, lo que indica un consumo estable a finales de año en lugar de un aumento estacional agresivo. Ecuador siguió siendo el principal proveedor, con 51,690 toneladas (+14% intermensual; +13% interanual), manteniendo su dominio estructural. India suministró 15,210 toneladas (+26% intermensual; +14% interanual), en consonancia con la reorientación en curso de los volúmenes indios hacia China, en medio de la incertidumbre provocada por los aranceles estadounidenses y la gestión de riesgos en otros mercados. Tailandia exportó 3,320 toneladas (-25% intermensual; -1% interanual), mientras que Indonesia aumentó a 1,890 toneladas (+111% intermensual; +66% interanual) desde una base más pequeña, lo que sugiere compras oportunistas de cobertura cuando la disponibilidad se alineó.
Las importaciones de Monodon disminuyeron desde los máximos de noviembre, pero se mantuvieron elevadas en comparación con el año pasado, alcanzando las 7590 toneladas en la Bolsa de Londres (-23% intermensual; +25% interanual). India se mantuvo como el principal impulsor con 7000 toneladas (-20% intermensual; +40% interanual), lo que refuerza la posición de China como el principal mercado de crecimiento global para el Monodon, a diferencia de otros mercados importantes donde la demanda sigue siendo estructuralmente limitada.
Perspectivas
El comercio de China a finales del año anterior indica un mercado que está absorbiendo volumen selectivamente, priorizando la cobertura sobre la acumulación activa de inventario. Los valores de importación
REPORTE DE MERCADO
continúan superando los volúmenes estructuralmente, respaldados por las elevadas estructuras de costos en las fases iniciales, incluyendo precios en origen, procesamiento y logística.
A principios de 2026, se espera que las compras se mantengan disciplinadas, con la acumulación de inventario probablemente concentrada en enero, antes del pico de
demanda del Año Nuevo Lunar a mediados de febrero de 2026.
Sin embargo, la sensibilidad a los precios sigue siendo alta. Cualquier nueva presión alcista por parte de Ecuador, el principal proveedor de China, probablemente limitaría el apetito de los importadores y mantendría las compras tácticas.
Al mismo tiempo, China debería seguir siendo un mercado de equilibrio clave para la oferta reasignada en medio de las presiones arancelarias actuales en los EE. UU., apoyando así la fortaleza continua de los volúmenes indios y manteniendo el impulso relativo de China en Monodon en relación con otros destinos•
Importaciones de China en 2023, 2024 y 2025 (volúmenes y precio promedio/kg)
Volumen de importación de Vannamei al mercado chino (2023, 2024 y 2025)
Importación de camarón de Estados Unidos
Autores:
Jim Kenny jkenny@urnerbarry.com
Ángel Rubio angel.rubio@expanamarkets.com
Urner Barry
Las importaciones de camarón de EE. UU. ascendieron a 136,6 millones de libras en noviembre, lo que supone un descenso interanual del 14%, pero un aumento del 5,5% con respecto a octubre. El arancel del 50% impuesto a la India, en vigor desde octubre, provocó una fuerte caída interanual del 32% en los envíos indios, mientras que Ecuador aumentó un 56% para capturar aproximadamente un tercio del mercado estadounidense, superando con creces a la India. Se espera que este cambio persista al menos durante diciembre y enero, incluso aunque el arancel de la India se haya reducido ahora al 18%.
Los volúmenes de Indonesia cayeron un 49% interanual, pero aumentaron un 13% con respecto a octubre, lo que sugiere una modesta recuperación a corto plazo a pesar de los continuos obstáculos arancelarios y normativos. Vietnam registró un aumento interanual del 7%, aunque su cuota global sigue siendo modesta. Los camarones pelados siguieron siendo la categoría dominante, con 67 millones de libras, un 9% menos que el año anterior, pero un 14% más que en octubre, lo que subraya el liderazgo actual de Ecuador. Los camarones con cáscara (-13%), cocidos (-22%) y rebozados (-16%) se sitúan por debajo de los niveles históricos.
Los costos de reposición aumentaron en todos los segmentos, liderados por los pelados (+8% hasta 4,17 $/lb) y los empanizados (+12% hasta 3,25 $/lb). Se espera que la oferta siga siendo escasa, con los importadores cada vez más inclinados hacia Ecuador y una demanda constante de formatos de valor añadido, ya que persisten las restricciones de suministro en Asia.
Por país
India: Las importaciones estadounidenses procedentes de la India en noviembre ascendieron a 40,7 millones de libras, lo que refleja los efectos continuados del arancel del 50% impuesto en octubre. Aunque los volúmenes se mantuvieron cercanos a las previsiones de los exportadores, los flujos siguen estando muy por debajo de los niveles del año anterior. La reciente reducción del arancel al 18% debería favorecer los envíos futuros, pero las llegadas durante el invierno reflejan en gran medida los contratos celebrados bajo el régimen arancelario más elevado.
Ecuador: El país siguió registrando fuertes volúmenes en noviembre, con 46,0 millones de libras, manteniendo su posición dominante en el sector del camarón pelado y compensando algunas carencias asiáticas. La capacidad de sustitución de Ecuador es limitada en los formatos cocidos, donde el suministro asiático sigue siendo líder. Las previsiones de exportación indican que Ecuador seguirá siendo un motor clave en el sector del camarón pelado, pero su impacto en los segmentos cocido y de valor añadido es comparativamente modesto.
Indonesia: Las importaciones procedentes de Indonesia disminuyeron hasta los 16,3
REPORTE DE MERCADO
millones de libras en noviembre, tras los mayores volúmenes registrados en octubre.
El suministro sigue estando por debajo de la media debido a las anteriores perturbaciones normativas y a las restricciones arancelarias. Las perspectivas de recuperación dependen de la estabilidad normativa y de los cambios en la demanda estadounidense, y la mayor parte del impacto a corto plazo está vinculado a las futuras llegadas en virtud de cualquier condición comercial ajustada.
Vietnam: Las importaciones procedentes de Vietnam alcanzaron los 17,7 millones de libras en noviembre, siguiendo de cerca los patrones históricos. La demanda se mantiene estable pero modesta, y los aranceles y la menor compra de EE. UU. limitan el movimiento al alza. Las perspectivas basadas en las exportaciones apuntan a flujos estables, aunque sin un aumento sustancial, ya que persisten las condiciones del mercado.
Exportaciones de camarones
Las exportaciones de Ecuador aumentaron un 4% interanual en noviembre, impulsadas por un sólido crecimiento hacia EE. UU. (+25%) y Europa (+35%), mientras que los envíos a China disminuyeron ligeramente. En lo que va de año, Ecuador ha registrado un aumento del 14%, lo que ilustra un claro giro hacia los mercados occidentales
a medida que se fortalece la demanda de importaciones de EE. UU.
Las exportaciones de la India aumentaron un 11%, destacando el aumento hacia China (+283%), impulsado por las compras estacionales, y hacia Europa (+88%), tras su nuevo acuerdo comercial con la UE. Los volúmenes destinados a EE. UU. cayeron un 21%, ya que los aranceles redirigieron los flujos y fomentaron un reajuste estratégico hacia mercados no estadounidenses.
Las exportaciones de Indonesia aumentaron un 30% en diciembre, principalmente debido a la mayor demanda asiática, aunque los envíos a Estados Unidos descendieron un 28%. Los efectos de la normativa y los aranceles siguen determinando la composición de las exportaciones de Indonesia.
En general, el cambio más importante ha sido el rápido giro de la India hacia China y Europa, mientras que el aumento de los volúmenes de Ecuador hacia Estados Unidos y la UE pone de relieve los ajustes más amplios de la oferta en respuesta a la dinámica del comercio mundial y los aranceles.
Importaciones con cáscara
Las importaciones con cáscara disminuyeron un 13% interanual en noviembre, con un total de 36,7 millones de libras, por debajo
de la media de los últimos tres años, debido a la moderada demanda estacional. Ecuador aumentó su cuota en un 56% y ahora representa más de la mitad del total de las llegadas. Los volúmenes de la India disminuyeron un 40%, a pesar de la reducción de los aranceles (ahora en el 18%), lo que favorece unos costos de sustitución más competitivos en el futuro. Los volúmenes de Indonesia cayeron un 76%, aunque se está produciendo una recuperación parcial tras las perturbaciones normativas. México y Argentina registraron aumentos, lo que contribuyó a una mayor diversificación del suministro.
Por tamaño, los camarones más grandes (<15) disminuyeron un 26% interanual, mientras que los de tamaño medio (16-30) se mantuvieron en su mayoría estables o al alza, con un aumento del 11% en las de 21-25 y del 5% en las de 26-30. Los de tamaño más pequeño (31-70) registraron un descenso de dos dígitos, lo que refleja la moderación de la actividad al contado y la limitada demanda después de la cosecha. Los costos de reposición subieron un 4,7% interanual hasta alcanzar los 4,02 $/lb, con un tono estable o firme liderado por Ecuador y Latinoamérica. Dada la persistencia de los contratos de suministro de Asia y la incertidumbre regulatoria, los compradores estadounidenses se mantienen cautelosos y gestionan estrictamente sus existencias
de cara a principios de 2026. La reducción de los aranceles de la India podría aliviar gradualmente la presión sobre los precios, mientras que la recuperación de Indonesia sigue siendo incompleta.
Precios con cáscara
Los precios con cáscara se mantienen estables o ligeramente a la baja en febrero, con los índices de referencia de Expana en 4,25 $/lb para HLSO y 4,45 $/lb para 2125 unidades. Los costos de reposición en el extranjero aumentaron en noviembre, alcanzando los 3,85 $/lb (Ecuador) y 4,43 $/ lb con aranceles, lo que redujo la diferencia con respecto a los índices de referencia de EE. UU.
Para los tamaños 21-25, los costos de reposición se sitúan en 3,47 $/lb (3,99 $/lb con aranceles), frente a los 4,45 $/ lb de Expana. Para los tamaños 41-50, la reposición en noviembre se situó en 2,56 $/lb (o 2,94 $/lb con aranceles), mientras que el índice de referencia de Expana es ahora de 3,60 $/lb. La diferencia de precios se ha reducido, lo que limita una mayor caída y señala un menor margen para los descuentos.
La confianza del mercado se mantiene estable o débil, con los compradores centrados en la gestión de inventarios y posiciones a futuro cautelosas. La reducción de los aranceles de la India está aliviando los costos de desembarque, pero se espera que los firmes valores de sustitución de América Latina y la recuperación incompleta de Asia impulsen la dirección del mercado a principios de 2026.
Importaciones de camarón pelado
Las importaciones de camarón pelado disminuyeron un 9% interanual en noviembre, hasta alcanzar un total de 66,9 millones de libras, lo que supone un descenso con respecto a los máximos de mediados de año, pero sigue estando por encima de la media estacional. Los volúmenes acumulados en lo que va de año han aumentado un 7,5%, lo que refleja la resistencia de la demanda y las compras anticipadas.
La cuota de la India se redujo al 40% en noviembre, ya que las llegadas se
mantuvieron por debajo del arancel del 50%; Ecuador subió al 33%, superando con creces a la India como principal proveedor. Los volúmenes de Indonesia cayeron un 21%, pero muestran una recuperación parcial; la cuota de Vietnam bajó, mientras que Argentina registró ganancias en lo que va de año, pero sigue siendo un contribuyente menor.
La mayor presencia de Ecuador está estabilizando el suministro y compensando el déficit asiático, ya que los volúmenes tienden a bajar estacionalmente en el cuarto trimestre. Los costos de reposición han aumentado un 8% interanual (hasta 4,17 dólares/lb, sin aranceles), lo que sitúa los valores de desembarque cerca de los niveles de ventas actuales. La gestión de los costos y la flexibilidad en el abastecimiento siguen siendo fundamentales a medida que el mercado se acerca a 2026.
Precios del camarón pelado
La confianza del mercado en el camarón pelado es estable pero mixta, con desequilibrios en el suministro entre los distintos tamaños y formas del producto. Las existencias no son notablemente escasas ni abundantes, pero la disponibilidad selectiva por segmentos mantiene a los compradores centrados en los programas comerciales y el abastecimiento estratégico.
Los costos de reposición (líneas naranjas, aranceles incluidos) siguen siendo elevados tras los picos del cuarto trimestre, y los valores de desembarque de noviembre para la India y Ecuador se acercan a los precios de referencia de Expana. Los precios de referencia de febrero están por encima de los niveles del año anterior, y los márgenes más ajustados limitan nuevas caídas; los descuentos al contado persisten, pero son menos frecuentes. Ahora que los aranceles de la India son mucho más competitivos en comparación con los de Ecuador (18% frente a 15%), se espera que la demanda vuelva gradualmente a la India, aunque el alcance dependerá de las señales de precios y la disponibilidad emergente.
El segmento pelado sigue siendo volátil, ya que los participantes en el mercado observan el reajuste de las cadenas de suministro. La diferencia real entre los costos de reposición y los precios internos es menor de lo que
indican los gráficos, dado el aumento de los gastos de logística, almacenamiento y aduanas. Seguirá siendo esencial una gestión cuidadosa de las existencias y las compras, ya que las presiones sobre los costos y los desajustes en la disponibilidad persistirán hasta principios de 2026.
Importaciones de productos cocinados y empanizados
Las importaciones de camarones cocinados cayeron un 22% interanual en noviembre hasta alcanzar los 21,3 millones de libras, lo que sigue estando por debajo de los niveles anteriores a 2023. Los volúmenes acumulados en lo que va de año aumentaron un 6%, liderados por India (+27%) y Tailandia (+2%), mientras que Indonesia y Vietnam registraron un nuevo descenso debido a las presiones arancelarias y de costos. La demanda del programa se mantiene estable, pero la actividad puntual se ve limitada por el estrechamiento de los márgenes y el aumento de los precios en el extranjero.
Las importaciones de camarones empanizados descendieron un 16% interanual en noviembre, con un volumen en lo que va de año prácticamente plano (-0,4%). Indonesia registró un fuerte retroceso, mientras que Tailandia y Ecuador experimentaron fuertes ganancias desde bases bajas; Vietnam, India y China continuaron descendiendo. Persistió la debilidad estacional, agravada por el traspaso de los aranceles y el aumento de los costos de los insumos, lo que ejerció presión sobre los márgenes de desembarque.
En general, la recuperación de los volúmenes de camarones cocidos y empanizados sigue siendo moderada, ya que el aumento de los aranceles y los elevados costos de importación han restringido las compras oportunistas y han limitado el potencial alcista. Las estrategias de inventario siguen siendo cautelosas, centrándose en las necesidades básicas de los programas.
Precios de los productos cocidos y con valor añadido
Los precios de los camarones cocidos se mantienen estables, con los índices de referencia de Expana para febrero en 8,05 dólares/lb para los camarones cocidos y
REPORTE DE MERCADO
5,98 dólares/lb para los camarones con valor añadido. Los costos de reposición se dispararon en noviembre, lo que elevó los valores de desembarque de la India a 6,46 dólares/lb (con aranceles) y los de Indonesia a 5,63 dólares/lb, ambos igualando o superando los índices de referencia y reduciendo los márgenes. La mayoría de las existencias de EE. UU. siguen vinculadas a los recientes aumentos de los aranceles, lo que mantiene elevados los costos de importación hasta principios de 2026.
La actividad al contado sigue siendo moderada, ya que el suministro asiático es limitado. El negocio del programa domina, y los compradores se mantienen cautelosos en sus adquisiciones. El segmento de Indonesia está empezando a recuperarse, pero los continuos obstáculos de la FDA siguen limitando el volumen y la flexibilidad.
De cara al futuro, la reducción de los aranceles de la India al 18% debería contribuir a aliviar las presiones sobre los costos de desembarque con el tiempo, reforzando su papel como proveedor líder. La evolución de los precios dependerá de las tendencias de sustitución, el suministro regional y la demanda constante de programas. Los gastos de logística y financiación, excluidos de los gráficos de costos de sustitución, comprimen aún más los márgenes, lo que requiere estrategias
de inventario disciplinadas, ya que los cambios arancelarios y normativos siguen configurando el mercado.
Precios en origen
Los precios en origen de las materias primas aumentaron en la mayoría de los lugares de origen durante el cuarto trimestre de 2025. Los valores en origen de la India, ajustados por rendimiento y convertidos a USD/lb, incluidos los aranceles, superaron brevemente los precios de venta de HLSO estadounidense a finales de 2025, lo que redujo drásticamente los márgenes de los agricultores, los envasadores y los importadores estadounidenses por igual. En el caso del segmento PDTO, los costos en origen de la India se mantuvieron justo por debajo de los niveles de venta de referencia, pero lo suficientemente cerca como para limitar los rendimientos en toda la cadena. Los precios en origen de Indonesia se mantuvieron elevados, pero por debajo de los máximos de la India, mientras que los de Ecuador se suavizaron, lo que ofreció una ventaja relativa en cuanto a costos para los compradores estadounidenses a valor nominal.
Los gráficos ponen de relieve cómo el aumento de los costos de los insumos y los aranceles han reducido los márgenes en todas las etapas, con el mayor impacto en
el HLSO procedente de la India e Indonesia. Ecuador sigue siendo un origen más competitivo, ya que sus precios en origen bajaron mientras que los de otros países subieron. El abastecimiento estratégico y la gestión del rendimiento siguen siendo cruciales, ya que la volatilidad persiste.
Ahora que el arancel de la India se ha reducido al 18%, los costos de desembarque deberían disminuir para las futuras llegadas, lo que ayudará a restablecer los márgenes. Se prevé que continúen las variaciones según el origen, con cambios en las ventajas de costos en función de los cambios del mercado y de la normativa.
Descargo de responsabilidad: Los precios en origen se ajustaron únicamente por la pérdida de rendimiento, convirtiendo el HOSO en HLSO y el PDTO en un tamaño equivalente, utilizando factores de conversión estándar. Los gráficos son ilustrativos y no estiman los costos de procesamiento, logística u otros. Los precios se convirtieron de la moneda local por kg a USD por libra.
Camarón silvestre del Golfo
Los desembarques de camarones silvestres del Golfo se mantuvieron en mínimos históricos hasta septiembre de 2025, con un descenso interanual de más del 50% en los volúmenes acumulados, hasta solo 14,3
millones de libras. Las capturas estacionales se han mantenido muy por debajo de los niveles habituales, lo que pone de relieve la persistente escasez de suministro. La cuota nacional del Golfo en el suministro total de camarones de EE. UU. cayó hasta solo el 2% en lo que va de año, lo que supone un nuevo mínimo y refuerza el creciente dominio de las importaciones en el mercado estadounidense.
Los precios del camarón salvaje HLSO del Golfo (U15, marrón) se mantuvieron firmes y estables, con cotizaciones al contado cercanas a los 12 dólares por libra y cotizando con una prima notable respecto al producto de criadero e importado.
Los continuos desembarques anémicos y la reducción de la oferta han mantenido las posiciones del mercado del Golfo relativamente estables. Se espera que la sustitución por importaciones persista, ya que la disponibilidad de camarón salvaje nacional se mantiene cerca de mínimos históricos.
Camarones al por menor
Las oportunidades de compra de camarones al por menor cayeron a solo 46 000 en las primeras seis semanas de 2026, la cifra más baja registrada. Los precios de los productos destacados subieron a 8,43 dólares por libra, lo que supone un máximo en 5 años y pone de relieve la continua inflación de los costos, junto con la disminución de las actividades promocionales.
Las ventas minoristas totales de camarones aumentaron un 1,1% interanual durante las primeras cuatro semanas, con un aumento del 2,6% en las ventas de camarones congelados y del 1,4% en las ventas de camarones frescos. Las ventas en dólares se ven impulsadas en gran medida por el aumento de los precios, mientras que la demanda subyacente sigue siendo moderada.
Los minoristas están reduciendo los descuentos agresivos debido a la persistencia de las presiones de los costos por el lado de la oferta, lo que limita el crecimiento del volumen a pesar de los precios elevados y los mínimos históricos en la actividad promocional. Los riesgos para el consumo
siguen siendo pronunciados a medida que aumenta la resistencia a los precios.
Otros factores impulsores Divisas
La competitividad del mercado del camarón sigue siendo sensible a las fluctuaciones monetarias, ya que la rupia india, la rupia indonesia y el dong vietnamita se han debilitado notablemente frente al dólar estadounidense desde principios de 2024. Esta depreciación hace que las exportaciones de camarones de estos importantes proveedores sean más atractivas en términos de dólares, lo que compensa en parte el aumento de los costos de producción y de reposición. Sin embargo, la debilidad actual también podría indicar retos macroeconómicos más amplios, como un acceso más restringido a la liquidez en dólares, lo que podría afectar a la adquisición de materias primas y a los flujos comerciales internacionales. Para los importadores estadounidenses, una moneda más débil del proveedor suele reducir los costos de desembarque, pero la ventaja se ve a veces contrarrestada por los altos aranceles y los elevados precios de los insumos. Es fundamental seguir de cerca las tendencias cambiarias para la adquisición de camarones a plazo.
Harina de pescado
Los precios de la harina de pescado se han recuperado considerablemente desde finales de 2025, y los grados chilenos y peruanos se acercan ahora a los 2200 USD/tm. Como ingrediente básico de la alimentación del camarón de acuicultura, el aumento de los costos de la harina de pescado incrementa directamente los costos de producción de los envasadores y los acuicultores, especialmente en Asia y América Latina. Esto añade presión al alza sobre los precios en origen y, a su vez, sobre los costos de reposición para los importadores estadounidenses. La volatilidad que se observa en el gráfico pone de relieve la sensibilidad de los mercados del camarón a las perturbaciones del suministro mundial de proteínas: cualquier aumento sostenido de los precios de los piensos puede traducirse en un aumento de los precios del camarón y en unos márgenes más ajustados en toda la cadena de suministro, lo que repercute tanto
en las negociaciones de precios como en la gestión de las existencias.
Harina de soja
La harina de soja es otro ingrediente clave en la alimentación del camarón, especialmente en los segmentos de cultivo de vannamei. El gráfico muestra descensos constantes de los precios en Argentina, Brasil y China desde mediados de 2023, lo que ha contribuido a moderar los costos de alimentación de los productores de camarón, incluso cuando la harina de pescado se ha recuperado. La bajada de los precios de la harina de soja puede aliviar la presión sobre los márgenes a nivel de la explotación, lo que favorece unos precios más competitivos del camarón para los compradores estadounidenses. Sin embargo, la continua volatilidad del suministro mundial de soja y de las compras asiáticas podría revertir estas ganancias. Para los importadores estadounidenses, es fundamental seguir la evolución de la harina de soja, ya que determina tanto los costos de producción como los precios de sustitución de los principales países exportadores.
Transporte y almacenamiento
Los costos de transporte y almacenamiento son fundamentales para los importadores de camarones estadounidenses y los proveedores extranjeros. Las tarifas de transporte marítimo desde la India, Indonesia y Vietnam se han normalizado hasta alcanzar los niveles previos a la pandemia, lo que ha aliviado las presiones de costos anteriores. Sin embargo, el almacenamiento y el depósito siguen aumentando, y el transporte refrigerado por carretera sigue siendo elevado. Si bien las tarifas marítimas estables favorecen unos costos de desembarque predecibles, el aumento del almacenamiento y el transporte nacional siguen ejerciendo presión sobre los márgenes y limitando el alivio de los precios. Con unos gastos logísticos por encima de los mínimos históricos, la gestión de los costos y la flexibilidad de la cadena de suministro son fundamentales para los compradores y exportadores de camarones, que se enfrentan a la inflación y a una demanda algo estable•
CNA realiza gestiones comerciales para atender restricciones puntuales en China
La Cámara Nacional de Acuacultura (CNA) mantiene activas y coordinadas gestiones técnicas, institucionales y diplomáticas para atender la suspensión temporal aplicada por la Administración General de Aduanas de China (GACC) a ocho empresas exportadoras de camarón ecuatoriano a finales de 2025. Esta medida se adoptó tras identificarse niveles de metabisulfito superiores a los límites permitidos durante controles realizados por la autoridad china, observaciones de carácter técnico que no estuvieron asociadas a riesgos sanitarios para los consumidores.
Desde el inicio de la situación, la CNA ha sostenido conversaciones directas con el Embajador de la República Popular China en Ecuador, con el objetivo de exponer el contexto técnico del sector y reforzar los canales de diálogo institucional. De manera paralela, ha dado seguimiento permanente a la Subsecretaría de Calidad e Inocuidad, coordinando el levantamiento de información sobre las acciones correctivas implementadas por los establecimientos involucrados, insumo clave para la elaboración de los Informes de Gestión de Crisis que deben ser presentados ante la GACC para el levantamiento de las suspensiones.
Ronda de negociación entre Ecuador y EE. UU. para reducir aranceles recíprocos
El Ministerio de Producción, Comercio Exterior e Inversiones (MPCEI) mantiene negociaciones con la Oficina del Representante Comercial de los Estados Unidos (USTR), con el objetivo de reducir los aranceles recíprocos aplicados a los productos ecuatorianos. El avance del proceso fue informado durante una reunión virtual realizada el viernes 30 de enero, en la que el ministro Luis Alberto Jaramillo y la viceministra de Comercio Exterior, Alexia Alcívar, expusieron el estado actual y los siguientes pasos de la negociación entre Ecuador y EE. UU.
Durante el encuentro se precisó que el acuerdo será gestionado directamente entre el MPCEI y la USTR, sin una firma intergubernamental. De igual manera, se anunció que en las próximas
Adicionalmente, la CNA ha acompañado y brindado información solicitada por parte de los establecimientos suspendidos, reforzando la importancia del control y dosificación de metabisulfito de sodio durante su cadena de proceso productivo, en línea con los requisitos del mercado chino. Asimismo, se ha realizado un seguimiento continuo a la situación de los contenedores y a la visibilidad de los establecimientos en el sistema CIFER, en articulación con la autoridad sanitaria ecuatoriana, con el propósito de facilitar y acelerar el proceso de normalización de las exportaciones.
El ministro de Producción, Comercio Exterior e Inversiones, Luis Jaramillo, participó en una mesa de trabajo con José Antonio Camposano, Presidente Ejecutivo de la CNA, en el marco de una reunión de trabajo realizada en 2025.
semanas se desarrollará una nueva ronda de negociación enfocada en cerrar los puntos aún pendientes y en la posible inclusión de productos en los anexos del acuerdo. Concluida esta etapa, se prevé avanzar hacia la suscripción del instrumento, en un contexto en el que otros países de la región ya han concluido procesos similares o se encuentran en su fase final•
Ecuador impulsa ante la OMSA una propuesta técnica para reducir barreras sanitarias al camarón
Con el fin de facilitar el acceso del camarón ecuatoriano a los mercados internacionales y reducir barreras sanitarias, se impulsó una gestión técnica ante la Organización Mundial de Sanidad Animal (OMSA) para adecuar los criterios sanitarios aplicables a productos destinados al consumo humano. La iniciativa responde a normativas que no consideran las particularidades del modelo productivo ecuatoriano ni el menor riesgo sanitario del camarón congelado en comparación con los organismos acuáticos vivos.
La propuesta plantea diferenciar normativamente el camarón congelado de los organismos vivos para reducir riesgos comerciales y fortalecer la competitividad del principal producto de exportación no petrolero del país. En este proceso, la Comisión Acuática de la OMSA tiene previsto analizar el 11 de febrero las observaciones presentadas
Coordinación con autoridades para una aplicación viable de nuevos controles
En diciembre pasado, la CNA gestionó una reunión de trabajo con el Ministerio del Interior para analizar la implementación de nuevos controles al metabisulfito de sodio, tras el anuncio oficial de medidas orientadas a evitar su uso indebido. El sector explicó que este insumo es de uso legítimo y seguro en la industria camaronera, donde se emplea como preservante y cuenta con sistemas de trazabilidad plenamente operativos.
En este sentido, se mantiene un seguimiento público y privado permanente para coordinar una aplicación técnica, ordenada y viable de los nuevos controles, garantizando el cumplimiento de la normativa, la prevención de desvíos ilícitos y evitando impactos innecesarios en la operatividad y competitividad del sector camaronero.
por Ecuador y otros países; mientras tanto, se mantienen reuniones de coordinación con aliados estratégicos para fortalecer la posición técnica nacional y respaldar una gestión orientada a la defensa y sostenibilidad del sector camaronero.
CNA solicitó revisión prioritaria del Impuesto a la Salida de Divisas
La CNA gestionó ante el Ministerio de Producción, Comercio Exterior e Inversiones la revisión prioritaria de las subpartidas arancelarias correspondientes a materias primas esenciales para el sector camaronero, como la soya y el trigo, insumos clave para la elaboración de alimento balanceado, dentro del marco de la revisión de las tarifas del Impuesto a la Salida de Divisas (ISD) para 2026.
Esta gestión busca evitar incrementos de tributos que afecten la estructura de costos del sector, considerando que el alimento balanceado representa cerca del 60% del valor total de producción del camarón. La CNA destacó ante las autoridades la importancia estratégica de la industria camaronera para la economía nacional, el empleo y las exportaciones, reiterando la necesidad de mantener condiciones tributarias que garanticen su continuidad productiva y competitividad internacional.
Análisis del Proyecto de Ley Orgánica para el Fortalecimiento de los Sectores Estratégicos de Minería y Energía
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura (CNA) y Presidente del Directorio de CORDEX, compareció ante la Comisión de Desarrollo Económico, Productivo y la Microempresa de la Asamblea Nacional, en el marco del análisis del Proyecto de Ley Orgánica para el Fortalecimiento de los Sectores Estratégicos de Minería y Energía, calificado como urgente en materia económica.
En la mesa de trabajo se planteó la visión del sector productivoexportador, destacando la importancia de esta iniciativa para garantizar energía confiable que permita sostener los procesos de tecnificación, competitividad internacional y generación de empleo.
“El desarrollo de fuentes de energía confiables y sostenibles es indispensable para continuar con los procesos de tecnificación,
NOTICIA EMPRESARIAL
Vitapro canaliza donación de arroz a más de 13 mil personas, a través del Banco de Alimentos Diakonía
Vitapro, en alianza con el Banco de Alimentos Diakonía, realizó la donación de 400 quintales de arroz a diversas comunidades beneficiarias e instituciones sociales, alcanzando a 13.300 personas. Como primera parte de la iniciativa, donó el producto a 47 familias de la comunidad Jesús del Gran Poder, ubicada en las cercanías a Milagro.
Esta acción se desarrolla en el marco de un proyecto impulsado por el Ministerio de Agricultura y Ganadería, orientado a evitar la pérdida de productos agrícolas ante el exceso de producción, al tiempo que se fortalece la seguridad alimentaria de poblaciones vulnerables mediante mecanismos de articulación entre el sector público, el sector privado y la sociedad civil. En este contexto, diversas empresas privadas se sumaron a la propuesta mediante la compra del arroz, permitiendo su posterior distribución social.
José Antonio Camposano, Presidente Ejecutivo de la Cámara Nacional de Acuacultura (CNA) y Presidente del Directorio de CORDEX, compareció ante la Comisión de Desarrollo Económico, Productivo y la Microempresa de la Asamblea Nacional
mantener la competitividad internacional y seguir generando empleo formal en el país”, puntualizó José Antonio Camposano.
En esta primera instancia, la Comisión continuará receptando los aportes de actores públicos y privados, con el fin de recoger observaciones, ajustar el texto y elaborar el informe para primer debate ante el Pleno de la Asamblea Nacional.
“Esta acción responde a la visión de Vitapro de generar valor compartido en los territorios donde opera, integrando su gestión productiva con acciones sociales que aporten al bienestar de las comunidades y al desarrollo del país. Desde esta perspectiva, la compañía concibe la sostenibilidad como un eje transversal, promoviendo así alianzas que permiten construir soluciones de impacto positivo y de largo plazo”, asegura Paúl Simball, jefe de Desarrollo Sostenible de la empresa.