GENOMES 5
T. A. Brown
Cover designed by Matthew McClements
Fifth edition published 2023 by CRC Press
6000 Broken Sound Parkway NW, Suite 300, Boca Raton, FL 33487-2742 and by CRC Press
4 Park Square, Milton Park, Abingdon, Oxon, OX14 4RN
CRC Press is an imprint of Taylor & Francis Group, LLC
© 2023 Terry Brown
Reasonable efforts have been made to publish reliable data and information, but the author and publisher cannot assume responsibility for the validity of all materials or the consequences of their use. The authors and publishers have attempted to trace the copyright holders of all material reproduced in this publication and apologize to copyright holders if permission to publish in this form has not been obtained. If any copyright material has not been acknowledged please write and let us know so we may rectify in any future reprint.
Except as permitted under U.S. Copyright Law, no part of this book may be reprinted, reproduced, transmitted, or utilized in any form by any electronic, mechanical, or other means, now known or hereafter invented, including photocopying, microfilming, and recording, or in any information storage or retrieval system, without written permission from the publishers.
For permission to photocopy or use material electronically from this work, access www.copyright.com or contact the Copyright Clearance Center, Inc. (CCC), 222 Rosewood Drive, Danvers, MA 01923, 978-750-8400. For works that are not available on CCC please contact mpkbookspermissions@tandf.co.uk
Trademark notice: Product or corporate names may be trademarks or registered trademarks and are used only for identification and explanation without intent to infringe.
Library of Congress Cataloging‑in‑Publication Data
Names: Brown, T. A. (Terence A.) author.
Title: Genomes 5 / T.A. Brown.
Other titles: Genomes five
Description: Fifth edition. | Boca Raton, FL : CRC Press, 2023. | Includes bibliographical references and index.
Identifiers: LCCN 2022058795 (print) | LCCN 2022058796 (ebook) | ISBN 9780367678661 (hbk) | ISBN 9780367674076 (pbk) | ISBN 9781003133162 (ebk)
Subjects: LCSH: Genomes.
Classification: LCC QH447 .B76 2023 (print) | LCC QH447 (ebook) | DDC 572.8/6--dc23/eng/20221209
LC record available at https://lccn.loc.gov/2022058795
LC ebook record available at https://lccn.loc.gov/2022058796
ISBN: 978-0-367-67866-1 (hbk)
ISBN: 978-0-367-67407-6 (pbk)
ISBN: 978-1-003-13316-2 (ebk)
DOI: 10.1201/9781003133162
Typeset in Utopia by Deanta Global Publishing Services, Chennai, India
Access the Instructor Resources by visiting www.routledge.com/9780367674076 and then clicking ‘Support Materials’, whereupon you can register as qualified instructor in order to gain access
Partial linkage is explained by the behavior of
3.4 LINKAGE ANALYSIS WITH DIFFERENT
analysis when planned breeding experiments are possible
mapping by human pedigree analysis
3.5 PHYSICAL MAPPING BY DIRECT
Conventional restriction mapping is only applicable to small DNA molecules
Optical mapping can locate restriction sites in longer DNA molecules
mapping with fluorescent probes
Further innovations extend the scope of optical mapping
3.6 PHYSICAL MAPPING BY ASSIGNING MARKERS TO DNA FRAGMENTS
Any unique sequence can be used as an STS
DNA fragments for STS mapping can be obtained as radiation hybrids
A clone library can be used as the mapping reagent
Many prokaryotic genomes have
4.1 METHODOLOGY
sequencing is the most popular short-
variety of other short-read sequencing methods have been devised
Nanopore sequencing is currently the longest long-read method
The Human Genome Project – genome sequencing in the heroic age 102
The human genome – genome sequencing in the modern age 104
The Neanderthal genome – assembly of an extinct genome using the human sequence as a reference 106
human genome – new challenges
5.1 GENOME ANNOTATION BY COMPUTER ANALYSIS OF THE DNA SEQUENCE 113
The coding regions of genes are open reading frames 113
Simple ORF scans are less effective with genomes of higher eukaryotes 114
Locating genes for noncoding RNA 116 Homology searches and comparative genomics give an extra dimension to gene prediction 117
5.2 GENOME ANNOTATION BY ANALYSIS OF GENE TRANSCRIPTS 119
Hybridization tests can determine if a fragment contains one or more genes 119 Methods are available for precise mapping of the ends of transcripts 120
Exon–intron boundaries can also be located with precision 121
5.3 ANNOTATION BY GENOME-WIDE RNA MAPPING 121
Tiling arrays enable transcripts to be mapped on to chromosomes or entire genomes 122
Transcript sequences can be directly mapped onto a genome 123
Obtaining transcript sequences by SAGE and CAGE 125
5.4 GENOME
PART 2 GENOME ANATOMIES
7
Ch APTER 6
IDENTIFYING GENE FUNCTIONS 131
6.1 COMPUTER ANALYSIS OF GENE FUNCTION 131
Homology reflects evolutionary relationships 131
Homology analysis can provide information on the function of a gene 132
Identification of protein domains can help to assign function to an unknown gene 133
Annotation of gene function requires a common terminology 134
6.2 ASSIGNING FUNCTION BY GENE INACTIVATION AND OVEREXPRESSION
Functional analysis by gene inactivation 136
Gene inactivation by genome editing 136
Gene inactivation by homologous recombination 137
Gene inactivation by transposon tagging and RNA interference 138
Gene overexpression can also be used to assess function 139
The phenotypic effect of gene inactivation or overexpression may be difficult to discern 140
6.3 UNDERSTANDING GENE FUNCTION BY STUDIES OF ITS EXPRESSION PATTERN AND PROTEIN PRODUCT
Reporter genes and immunocytochemistry can be used to locate where and when genes are expressed 142
CRISPR can be used to make specific changes in a gene and the protein it encodes 143
Other methods for site-directed mutagenesis 145
6.4 USING CONVENTIONAL GENETIC
7.1 NUCLEAR GENOMES ARE CONTAINED IN CHROMOSOMES
are made of DNA and protein
7.3 THE REPETITIVE DNA CONTENT OF
Ch APTER 8
8.1 THE PHYSICAL FEATURES OF
8.2 THE GENETIC FEATURES
Gene organization in the E. coli K12 genome 180
Operons are characteristic features of prokaryotic genomes 182
Prokaryotic genome sizes and gene numbers vary according to biological complexity 184
Genome sizes and gene numbers vary within individual species 185
Distinctions between prokaryotic species are further blurred by horizontal gene transfer 186
Metagenomes describe the members of a community 188
8.3 EUKARYOTIC ORGANELLE GENOMES
The endosymbiont theory explains the origin of organelle genomes 190
The physical and genetic features of organelle genomes 191
APTER 10
Ch APTER 9
9.1 THE GENOMES OF BACTERIOPHAGES AND EUKARYOTIC VIRUSES
Bacteriophage genomes have diverse structures and organizations 199
Replication strategies for bacteriophage genomes 201
Structures and replication strategies for eukaryotic viral genomes 202
Some retroviruses cause cancer 204
Genomes at the edge of life 205
9.2 MOBILE GENETIC ELEMENTS
RNA transposons with long terminal repeats are related to viral retroelements 206
Some RNA transposons lack LTRs 208
DNA transposons are common in prokaryotic genomes
DNA transposons are less common in eukaryotic genomes
Ch APTER 11
NMR spectroscopy is used to study the structures of small proteins
Gel retardation identifies DNA fragments that bind to proteins
Protection assays pinpoint binding sites with greater accuracy
Modification interference identifies nucleotides central to protein binding 244
Genome-wide scans for protein attachment sites 245
11.2 THE SPECIAL FEATURES OF DNA-BINDING PROTEINS
The helix–turn–helix motif is present in prokaryotic and eukaryotic proteins
Zinc fingers are common in eukaryotic proteins
Other nucleic acid-binding motifs
11.3 THE INTERACTION BETWEEN DNA AND ITS BINDING PROTEINS 249
Contacts between DNA and proteins
Direct readout of the nucleotide sequence
The conformation of the helix also influences protein binding
Ch APTER 12 TRANSCRIPTOMES
12.1 THE COMPONENTS OF THE TRANSCRIPTOME
The mRNA fraction of a transcriptome is small but complex
Short noncoding RNAs have diverse functions 258 Long noncoding RNAs are enigmatic transcripts 260
12.2 TRANSCRIPTOMICS: CATALOGING THE TRANSCRIPTOMES OF CELLS AND TISSUES
Microarray analysis and RNA sequencing are used to study the contents of transcriptomes
Single-cell studies add greater precision to transcriptomics
Spatial transcriptomics enables transcripts to be mapped directly in tissues and cells
12.3 SYNTHESIS OF THE COMPONENTS OF THE TRANSCRIPTOME
RNA polymerases are molecular machines for making
Transcription start-points are indicated by promoter
Synthesis of bacterial RNA is regulated by repressor and activator proteins
Synthesis of bacterial RNA is also regulated by control over transcription termination
Synthesis of eukaryotic RNA is regulated primarily by activator proteins 277
12.4 THE INFLUENCE OF RNA SPLICING ON THE COMPOSITION OF A TRANSCRIPTOME 280 The splicing pathway for eukaryotic pre-mRNA introns
The splicing process must have a high degree of precision
Enhancer and silencer elements specify alternative splicing pathways 284
Backsplicing gives rise to circular RNAs
12.5 THE INFLUENCE OF CHEMICAL MODIFICATION ON THE COMPOSITION OF A TRANSCRIPTOME
RNA editing alters the coding properties of some transcripts
Chemical modifications that do not affect the sequence of an mRNA
12.6 DEGRADATION OF THE COMPONENTS OF THE TRANSCRIPTOME 290 Several processes are known for nonspecific RNA turnover 291
RNA silencing was first identified as a means of destroying invading viral RNA
MicroRNAs regulate genome expression by causing specific target mRNAs to be degraded
13.1 STUDYING THE COMPOSITION OF A PROTEOME
The separation stage of a protein profiling project
The identification stage of a protein profiling project
Comparing the compositions of two proteomes 305 Analytical protein arrays offer an alternative approach to protein profiling
13.2 IDENTIFYING PROTEINS THAT INTERACT WITH ONE ANOTHER
Identifying the components of multiprotein complexes
Identifying proteins with functional interactions
Protein interaction maps display the interactions within a proteome
13.3 SYNTHESIS AND DEGRADATION OF THE COMPONENTS OF THE PROTEOME 313
Ribosomes are molecular machines for making proteins 313
During stress, bacteria inactivate their ribosomes in order to downsize the proteome 316
Initiation factors mediate large-scale remodeling of eukaryotic proteomes 317
The translation of individual mRNAs can also be regulated 318
Degradation of the components of the proteome 320
13.4 THE INFLUENCE OF PROTEIN PROCESSING ON THE COMPOSITION OF THE PROTEOME
The amino acid sequence contains instructions for protein folding 321
Some proteins undergo proteolytic cleavage 324
Important changes in protein activity can be brought about by chemical modification 325
13.5 BEYOND THE PROTEOME
The metabolome is the complete set of metabolites present in a cell
Systems biology provides an integrated description of cellular activity
14.1 THE RESPONSE OF THE GENOME TO EXTERNAL SIGNALS
Signal transmission by import of the extracellular
Receptor
Some signal transduction pathways have few steps between
Some
transduction pathways have many steps between
Some
transduction pathways operate via second messengers
Yeast mating types are determined by gene
Genome rearrangements are responsible for immunoglobulin and T-cell receptor diversities
Bacteriophage λ: a genetic switch enables a choice to be made between alternative
Bacillus sporulation: coordination of activities in two distinct cell types
Caenorhabditis elegans: the genetic basis to positional information and the determination of cell fate
flies: conversion of positional information into a segmented body plan
genes are universal features of
PART 4 HOW GENOMES
The Meselson–Stahl experiment proved that replication is
provide a solution to the
Origins of replication have been clearly defined in yeast 369
Origins in higher eukaryotes have been less easy to identify 370
15.3 EVENTS AT THE REPLICATION FORK 371
DNA polymerases are molecular machines for making (and degrading) DNA 371
DNA polymerases have limitations that complicate genome replication 373
Okazaki fragments must be joined together to complete lagging-strand replication 374
15.4 TERMINATION OF GENOME REPLICATION 376
Replication of the E. coli genome terminates within a defined region 376
Completion of genome replication 378
Telomerase completes replication of chromosomal DNA molecules, at least in some cells 380
Telomere length is implicated in cell senescence and cancer 382
Drosophila has a unique solution to the endshortening problem 383
15.5 REGULATION OF EUKARYOTIC GENOME REPLICATION 384
Genome replication must be synchronized with the cell cycle 384
Origin licensing is the prerequisite for passing the G1-S checkpoint 385
Replication origins do not all fire at the same time 386
The cell has various options if the genome is damaged 388
Bacteriophage λ uses site-specific recombination during the lysogenic infection cycle
Site-specific recombination is an aid in construction of genetically modified plants
16.1 HOMOLOGOUS RECOMBINATION
The Holliday and Meselson–Radding models for homologous recombination
The double-strand break model for homologous recombination
RecBCD is the most important pathway for homologous recombination in bacteria
E. coli has alternative pathways for homologous
excision repair is used to correct more extensive types of damage
repair corrects replication errors
and double-strand breaks can be repaired
Some types of damage can be repaired by homologous recombination
18.1 GENOMES: THE FIRST 10 BILLION YEARS 431
The first biochemical systems were centered on RNA 431
The first DNA genomes 433 How unique is life? 434
18.2 THE EVOLUTION OF INCREASINGLY COMPLEX GENOMES 436
Genome sequences provide extensive evidence of past gene duplications 436
A variety of processes could result in gene duplication 439
Whole-genome duplication is also possible 440
Smaller duplications can also be identified in the human genome and other genomes 443
Both prokaryotes and eukaryotes acquire genes from other species 444
Genome evolution also involves rearrangement of existing gene sequences 446
There are competing hypotheses for the origins of introns 448
The evolution of the epigenome 450
18.3 GENOMES: THE LAST 6 MILLION YEARS
The human genome is very similar to that of the chimpanzee 451
Paleogenomics is helping us understand the recent evolution of the human genome 453
18.4 GENOMES TODAY: DIVERSITY IN
HOW GENOMES ARE STUDIED
GENOMES, TRANSCRIPTOMES, AND PROTEOMES
Life as we know it is specified by the genomes of the myriad organisms with which we share the planet. Every organism possesses a genome that contains the biological information needed to construct and maintain a living example of that organism. Most genomes, including the human genome and those of all other cellular life forms, are made of DNA (deoxyribonucleic acid), but a few viruses have RNA (ribonucleic acid) genomes. DNA and RNA are polymeric molecules made up of chains of monomeric subunits called nucleotides. Each molecule of DNA comprises two polynucleotides wound around one another to form the famous double helix, the two strands held together by chemical bonds that link adjacent nucleotides into structures called base pairs The human genome, which is typical of the genomes of all multicellular animals, consists of two distinct parts (Figure 1.1):
• The nuclear genome comprises approximately 3,100,000,000 base pairs of DNA, divided into 24 linear molecules, the shortest 47,000,000 base pairs in length and the longest 249,000,000 base pairs, each contained in a different chromosome. These 24 chromosomes consist of 22 autosomes and the two sex chromosomes, X and Y. Altogether, some 44,500 genes are present in the human nuclear genome.
• The mitochondrial genome is a circular DNA molecule of 16,569 base pairs, up to ten copies of which are present in each of the energy-generating organelles called mitochondria. The human mitochondrial genome contains just 37 genes.
Each of the approximately 3 × 1013 cells in the adult human body has its own copy or copies of the nuclear genome, the only exceptions being those few cell types, such as red blood cells, that lack a nucleus in their fully differentiated state. The vast majority of cells are diploid and so have two copies of each autosome, plus two sex chromosomes, XX for females or XY for males – 46 chromosomes in all. These are called somatic cells, in contrast to sex cells, or gametes, which are haploid and have just 23 chromosomes, comprising one of each autosome and one sex chromosome. Each cell also has multiple copies of the mitochondrial genome, 2000–7000 copies in somatic cells such as those in the liver and heart tissue, and over 100,000 in each female oocyte.
1.1 DNA
1.2 RNA AND THE TRANSCRIPTOME
1.3 PROTEINS AND THE PROTEOME
The genome is a store of biological information, but on its own it is unable to release that information to the cell. Utilization of the biological information contained in the genome requires the coordinated activity of enzymes and other proteins, which participate in a complex series of biochemical reactions referred to as genome expression (Figure 1.2). The initial product of genome expression is the transcriptome, a collection of RNA molecules derived from those genes that are active in the cell at a particular time. The transcriptome is maintained by the process called transcription, in which individual genes are copied into RNA molecules. The second product of genome expression is the proteome, the cell’s repertoire of proteins, which specifies the nature of the biochemical reactions that the cell is able to carry out. The proteins that make up the proteome are synthesized by translation of some of the individual RNA molecules present in the transcriptome.
This book is about genomes and genome expression. It explains how genomes are studied (Part 1), how they are organized (Part 2), how they function (Part 3), and how they replicate and evolve (Part 4). It was not possible to write this book until quite recently. Since the 1950s, molecular biologists have studied individual genes or small groups of genes, and from these studies have built up a wealth of knowledge about how genes work. But only during the last few years have techniques been available that make it possible to examine entire genomes. Individual genes are still intensively studied, but information about individual genes is now interpreted within the context of the genome as a whole. This new, broader emphasis applies not just to genomes but to all of biochemistry and cell biology. No longer is it sufficient simply to understand individual biochemical pathways or subcellular processes. The challenge now is provided by systems biology, which attempts to link together these pathways and processes into networks that describe the overall functioning of living cells and living organisms.
This book will lead you through our knowledge of genomes and show you how this exciting area of research is underpinning our developing understanding of biological systems. First, however, we must pay attention to the basic principles of molecular biology by reviewing the key features of the three types of biological molecule involved in genomes and genome expression: DNA, RNA, and protein.
1.1 DNA
DNA was discovered in 1869 by Johann Friedrich Miescher, a Swiss biochemist working in Tübingen, Germany. The first extracts that Miescher made from human white blood cells were crude mixtures of DNA and chromosomal proteins, but the following year he moved to Basel, Switzerland (where the research
Human family
Human cell
Nuclear genomeMitochondrial genome
Figure 1.2 Genome expression. The genome specifies the transcriptome, and the transcriptome specifies the proteome.
Figure 1.1 The nuclear and mitochondrial components of the human genome.
institute named after him is now located), and prepared a pure sample of nucleic acid from salmon sperm. Miescher’s chemical tests showed that DNA is acidic and rich in phosphorus, and also suggested that the individual molecules are very large, although it was not until the 1930s, when biophysical techniques were applied to DNA, that the huge lengths of the polymeric chains were fully appreciated.
Genes are made of DNA
The fact that genes are made of DNA is so well known today that it can be difficult to appreciate that for the first 75 years after its discovery the true role of DNA was unsuspected. As early as 1903, W.S. Sutton had realized that the inheritance patterns of genes parallel the behavior of chromosomes during cell division, an observation that led to the chromosome theory, the proposal that genes are located in chromosomes. Examination of cells by cytochemistry, which makes use of stains that bind specifically to just one type of biochemical, showed that chromosomes are made of DNA and protein, in roughly equal amounts. Biologists at that time recognized that billions of different genes must exist and the genetic material must therefore be able to take many different forms. But this requirement appeared not to be satisfied by DNA, because in the early part of the twentieth century it was thought that all DNA molecules were the same. On the other hand, it was known, correctly, that proteins are highly variable, polymeric molecules, each one made up of a different combination of 20 chemically distinct, amino acid monomers (Section 1.3). Genes simply had to be made of protein, not DNA.
The errors in understanding DNA structure lingered on, but by the late 1930s it had become accepted that DNA, like protein, has immense variability. The notion that protein was the genetic material initially remained strong, but was eventually overturned by the results of two important experiments:
• Oswald Avery, Colin MacLeod, and Maclyn McCarty showed that DNA is the active component of the transforming principle, a bacterial cell extract which, when mixed with a harmless strain of Streptococcus pneumoniae, converts these bacteria into a virulent form capable of causing pneumonia when injected into mice (Figure 1.3A). In 1944, when the results of this experiment were published, only a few microbiologists appreciated that transformation involves transfer of genes from the cell extract into the living bacteria. However, once this point had been accepted, the true meaning of the ‘Avery experiment’ became clear: bacterial genes must be made of DNA.
• Alfred Hershey and Martha Chase used radiolabeling to show that, when a bacterial culture is infected with bacteriophages (also called phages, a type of virus), DNA is the major component of the bacteriophages that enters the cells (Figure 1.3B). This was a vital observation because it was known that, during the infection cycle, the genes of the infecting bacteriophages are used to direct synthesis of new bacteriophages, and this synthesis occurs within the bacteria. If it is only the DNA of the infecting bacteriophages that enters the cells, then it follows that the genes of these bacteriophages must be made of DNA.
Although from our perspective these two experiments provide the key results that tell us that genes are made of DNA, biologists at the time were not so easily convinced. Both experiments have limitations that leave room for skeptics to argue that protein could still be the genetic material. For example, there were worries about the specificity of the deoxyribonuclease enzyme that Avery and colleagues used to inactivate the transforming principle. This result, a central part of the evidence for the transforming principle being DNA, would be invalid if, as seemed possible, the enzyme contained trace amounts of a contaminating protease and hence was also able to degrade
Mouse survivesHarmless
Harmless bacteria + transforming principle
Mouse diesHarmless transforming principle treated with protease or ribonuclease
Harmless bacteria + transforming principle treated with deoxyribonuclease
Figure 1.3 The two experiments that suggested that genes are made of DNA. (A) Avery and colleagues showed that the transforming principle is made of DNA. The top two panels show what happens when mice are injected with harmless Streptococcus pneumoniae bacteria, with or without addition of the transforming principle, a cell extract obtained from a virulent strain of S. pneumoniae. When the transforming principle is present, the mouse dies, because the genes in the transforming principle convert the harmless bacteria into the virulent form, with these virulent bacteria subsequently being recovered from the lungs of the dead mouse. The lower two panels show that treatment with protease or ribonuclease has no effect on the transforming principle, but that the transforming principle is inactivated by deoxyribonuclease. (B) The Hershey–Chase experiment used T2 bacteriophages, each of which comprises a DNA molecule contained in a protein capsid attached to a ‘body’ and ‘legs’ that enable the bacteriophage to attach to the surface of a bacterium and inject its genes into the cell. The DNA of the bacteriophages was labeled with 32P, and the protein with 35S. A few minutes after infection, the culture was agitated to detach the empty bacteriophage particles from the cell surface. The culture was then centrifuged, which collects the bacteria plus bacteriophage genes as a pellet at the bottom of the tube, but leaves the lighter bacteriophage particles in suspension. Hershey and Chase found that the bacterial pellet contained 70% of the 32P-labeled component of the bacteriophages (the DNA) but only 20% of the 35S-labeled material (the bacteriophage protein). In a second experiment, Hershey and Chase showed that new bacteriophages produced at the end of the infection cycle contained less than 1% of the protein from the parent bacteriophages. For more details of the bacteriophage infection cycle, see Figure 2.27.
4Chapter 1: Genomes, transcriptomes, and proteomes protein. Neither is the bacteriophage experiment conclusive, as Hershey and Chase stressed when they published their results: ‘Our experiments show clearly that a physical separation of phage T2 into genetic and non-genetic parts is possible The chemical identification of the genetic part must wait, however, until some questions have been answered.’ In retrospect, these two experiments are important not because of what they tell us but because they alerted biologists to the fact that DNA might be the genetic material and was therefore worth studying. It was this that influenced Watson and Crick to work on DNA and, as we will see below, it was their discovery of the double-helix structure, which solved the puzzling question of how genes can replicate, which really convinced the scientific world that genes are made of DNA.
Mouse survives
Mouse dies
Agitate in blender
Centrifuge
Phage attached to bacteria
Pellet of bacteria
Virulent bacteria
Virulent bacteria
(A) The transforming principle
(B) The Hershey–Chase experiment
DNA is a polymer of nucleotides
The names of James Watson and Francis Crick are so closely linked with DNA that it is easy to forget that when they began their collaboration in October 1951 the detailed structure of the DNA polymer was already known. Their contribution was not to determine the structure of DNA per se, but to show that in living cells two DNA chains are intertwined to form the double helix. First, therefore, we should examine what Watson and Crick knew before they began their work. DNA is a linear, unbranched polymer in which the monomeric subunits are four chemically distinct nucleotides that can be linked together in any order in chains hundreds, thousands, or even millions of units in length. Each nucleotide in a DNA polymer is made up of three components (Figure 1.4):
• 2´-Deoxyribose, which is a pentose, a type of sugar composed of five carbon atoms. These five carbons are numbered 1´ (spoken as ‘one-prime’), 2´, and so on. The name ‘2´-deoxyribose’ indicates that this particular sugar is a derivative of ribose, in which the hydroxyl (–OH) group attached to the 2´-carbon of ribose has been replaced by a hydrogen (–H) group.
• A nitrogenous base, one of cytosine, thymine (single-ring pyrimidines), adenine, or guanine (double-ring purines). The base is attached to the 1´-carbon of the sugar by a β-N-glycosidic bond attached to nitrogen number one of the pyrimidine or number nine of the purine.
• A phosphate group comprising one, two, or three linked phosphate units attached to the 5´-carbon of the sugar. The phosphates are designated α, β, and γ, with the α-phosphate being the one directly attached to the sugar.
A molecule made up of just the sugar and base is called a nucleoside; the addition of the phosphates converts this into a nucleotide. Although cells contain nucleotides with one, two, or three phosphate groups, only the nucleoside triphosphates act as substrates for DNA synthesis. The full chemical names of the four nucleotides that polymerize to make DNA are:
• 2´-deoxyadenosine 5´-triphosphate
• 2´-deoxycytidine 5´-triphosphate
• 2´-deoxyguanosine 5-triphosphate
• 2´-deoxythymidine 5-triphosphate
The abbreviations of these four nucleotides are dATP, dCTP, dGTP, and dTTP, respectively, or when referring to a DNA sequence, A, C, G, and T, respectively.
(A) A nucleotide
(B) The four bases in DNA
Figure 1.4 The structure of a nucleotide. (A) The general structure of a deoxyribonucleotide, the type of nucleotide found in DNA. (B) The four bases that
Figure 1.5 A short DNA polynucleotide showing the structure of the phosphodiester bond. Note that the two ends of the polynucleotide are chemically distinct.
In a polynucleotide, individual nucleotides are linked together by phosphodiester bonds between their 5´- and 3´-carbons (Figure 1.5). From the structure of this linkage, we can see that the polymerization reaction (Figure 1.6) involves removal of the two outer phosphates (the β- and γ-phosphates) from one nucleotide and replacement of the hydroxyl group attached to the 3´-carbon of the second nucleotide. Note that the two ends of the polynucleotide are chemically distinct, one having an unreacted triphosphate group attached to the 5´-carbon (the 5´ or 5´-P terminus), and the other having an unreacted hydroxyl attached to the 3´-carbon (the 3´ or 3´-OH terminus). This means that the polynucleotide has a chemical direction, expressed as either 5´→3´ (down in Figure 1.5) or 3´→5´ (up in Figure 1.5). An important consequence of the polarity of the phosphodiester bond is that the chemical reaction needed to extend a DNA polymer in the 5´→3´ direction is different to that needed to make a 3´→5´ extension. The DNA polymerase enzymes present in living organisms are only able to carry out 5´→3´ synthesis, which adds significant complications to the process by which double-stranded DNA is replicated (Section 15.3).
The discovery of the double helix
In the years before 1950, various lines of evidence had shown that cellular DNA molecules are comprised of two or more polynucleotides assembled together in some way. The possibility that unraveling the nature of this assembly might provide insights into how genes work prompted Watson and Crick, among others, to try to solve the structure. According to Watson, in his book The Double Helix, their work was a desperate race against the famous American biochemist, Linus Pauling, who initially proposed an incorrect triple helix model, giving Watson and Crick the time they needed to complete the double-helix structure. It is now difficult to separate fact from fiction, especially regarding the part played by Rosalind Franklin, whose X-ray diffraction studies provided the bulk of the experimental data in support of the double helix and who was herself very close to solving the structure. The one thing that is clear is that the double helix, discovered by Watson and Crick on Saturday, March 7, 1953, was the single most important breakthrough in biology during the twentieth century.
The discovery of the double helix can be looked on as one of the first multidisciplinary biological research projects. Watson and Crick used four quite different types of information to deduce the double-helix structure:
• Biophysical data of various kinds were used to infer some of the key features of the structure. The water content of DNA fibers was particularly important because it enabled the density of the DNA in a fiber to be estimated. The number of strands in the helix and the spacing between the nucleotides
had to be compatible with the fiber density. Pauling’s triple helix model was based on an incorrect density measurement that suggested that the DNA molecule was more closely packed than is actually the case.
• X-ray diffraction patterns (Section 11.1), most of which were produced by Rosalind Franklin, revealed the detailed helical structure (Figure 1.7).
• The base ratios, which had been discovered by Erwin Chargaff of Columbia University, New York, enabled the pairing between the polynucleotides in the helix to be deduced. Chargaff had carried out a lengthy series of chromatographic studies of DNA samples from various sources and showed that, although the values are different in different organisms, the amount of adenine is always the same as the amount of thymine, and the amount of guanine equals the amount of cytosine (Figure 1.8). These base ratios led to the base-pairing rules, which were the key to the discovery of the double-helix structure.
• The construction of scale models of possible DNA structures, which was the only major technique that Watson and Crick performed themselves, enabled the relative positioning of the various atoms to be checked, to ensure that pairs that formed bonds were not too far apart, and that other atoms were not so close together as to interfere with one another.
Figure 1.6 The polymerization reaction that results in synthesis of a DNA polynucleotide. Synthesis occurs in the 5´→3´ direction, with the new nucleotide being added to the 3´-carbon at the end of the existing polynucleotide. The β- and γ-phosphates of the nucleotide are removed as a pyrophosphate molecule.
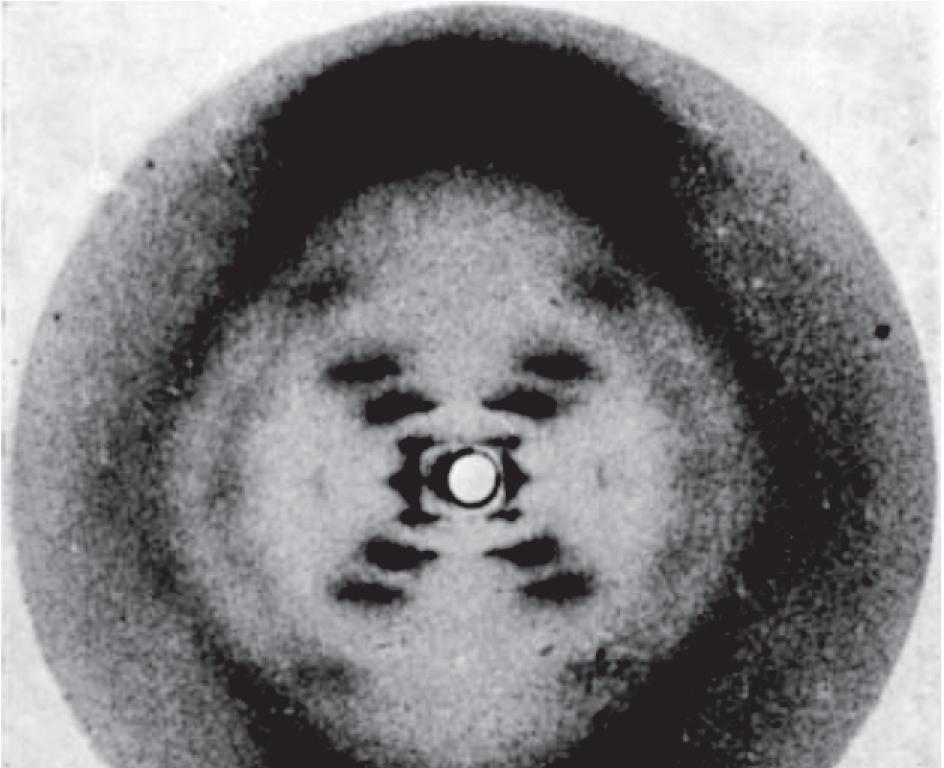

Figure 1.7 franklin’s ‘photo 51’ showing the X-ray diffraction pattern obtained with a fiber of DNA. The cross shape indicates that DNA has a helical structure, and the extent of the shadowing within the ‘diamond’ spaces above, below and to either side of the cross show that the sugar–phosphate backbone is on the outside of the helix (see Figure 1.9). The positions of the various smears that make up the arms of the cross enable dimensions such as the diameter, rise per base pair and pitch (see Table 1.1) of the molecule to be calculated. The ‘missing smears’ (the gap in each arm of the cross, marked by the arrows) indicate the relative positioning of the two polynucleotides. These missing smears enabled Watson and Crick to recognize that there are two grooves of different depths on the outer surface of the helix (see Figure 1.9). (From Franklin R & Gosling RG [1953] Nature 171:740-741. With permission from
Purify the DNA
Mild acid treatment breaks phosphodiester bonds
Chromatography to quantify each nucleotide
The double helix is stabilized by base-pairing and base-stacking
The double helix is right-handed, which means that if it were a spiral staircase and you were climbing upwards then the rail on the outside of the staircase would be on your right-hand side. The two strands run in opposite directions (Figure 1.9A). The helix is stabilized by two types of chemical interaction:
• Base-pairing between the two strands involves the formation of hydrogen bonds between an adenine on one strand and a thymine on the other strand, or between a cytosine and a guanine (Figure 1.9B). Hydrogen bonds are weak electrostatic interactions between an electronegative atom (such as oxygen or nitrogen) and a hydrogen atom attached to a second electronegative atom. Hydrogen bonds are longer than covalent bonds and are much weaker, typical bond energies being 8–29 kJ mol–1 at 25°C, compared with up to 348 kJ mol–1 for a single covalent bond between a pair of carbon atoms. As well as their role in the DNA double helix, hydrogen bonds stabilize protein secondary structures. The two base-pair combinations – A base-paired with T, and G base-paired with C – explain the base ratios discovered by Chargaff. These are the only pairs that are permissible, partly because of the geometries of the nucleotide bases and the relative positions of the atoms that are able to participate in hydrogen bonds, and partly because the pair must be between a purine

Figure 1.8 The base ratio experiments performed by Chargaff. DNA was extracted from various organisms and treated with acid to hydrolyze the phosphodiester bonds and release the individual nucleotides. Each nucleotide was then quantified by chromatography. The data show some of the actual results obtained by Chargaff. These indicate that, within experimental error, the amount of adenine is the same as that of thymine, and the amount of guanine is the same as that of cytosine.
Figure 1.9 The double-helix structure of DNA. (A) Two representations of the double helix. On the left, the structure is shown with the sugar–phosphate ‘backbones’ of each polynucleotide drawn as a gray ribbon with the base pairs in green. On the right the chemical structure for three base pairs is given. (B) A base-pairs with T, and G base-pairs with C. The bases are drawn in outline, with the hydrogen bonding indicated by dotted lines. Note that a G–C base pair has three hydrogen bonds whereas an A–T base pair has just two.
groove Sugar–phosphate backbone
Human cells Escherichia coli bacteria
5‘3‘ Base pair Hydrogen bonds
end
end
end
end
Adenine (A)Cytosine (C)Guanine (G )Thymine T)
and a pyrimidine: a purine–purine pair would be too big to fit within the helix, and a pyrimidine–pyrimidine pair would be too small.
• Base-stacking involves attractive forces between adjacent base pairs and adds stability to the double helix once the strands have been brought together by base-pairing. Base-stacking is sometimes called π–π interactions, because it is thought to involve the p electrons associated with the double bonds of the purine and pyrimidine structures. However, this hypothesis is now being questioned and the possibility that base-stacking involves a type of electrostatic interaction is being explored.
Both base-pairing and base-stacking are important in holding the two polynucleotides together, but base-pairing has added significance because of its biological implications. The limitation that A can only base-pair with T, and G can only base-pair with C, means that DNA replication can result in perfect copies of a parent molecule through the simple expedient of using the sequences of the preexisting strands to dictate the sequences of the new strands. This is template-dependent DNA synthesis, and it is the system used by all cellular DNA polymerases (Section 2.1). Base-pairing therefore enables DNA molecules to be replicated by a system that is so simple and elegant that as soon as the doublehelix structure was publicized by Watson and Crick, every biologist became convinced that genes really are made of DNA.
The double helix has structural flexibility
The double helix described by Watson and Crick, and shown in Figure 1.9A , is called the B-form of DNA or B-DNA. Its characteristic features lie in its dimensions: a helical diameter of 2.37 nm, a rise of 0.34 nm per base pair, and a pitch (the distance taken up by a complete turn of the helix) of 3.4 nm, corresponding to ten base pairs per turn. The DNA in living cells is thought to be predominantly in this B-form, but it is now clear that genomic DNA molecules are not entirely uniform in structure. This is mainly because each nucleotide in the helix has the flexibility to take up slightly different molecular shapes. To adopt these different conformations, the relative positions of the atoms in the nucleotide must change slightly. There are a number of possibilities but the most important conformational changes are:
• Rotation around the β-N-glycosidic bond, which changes the orientation of the base relative to the sugar: the two possibilities are called the anti and syn conformations (Figure 1.10A). Base rotation influences the positioning of the two polynucleotides.
• Sugar pucker, which refers to the three-dimensional shape of the sugar. The ribose component of the nucleotide does not have a planar structure: when viewed from the side, one or two of the carbon atoms are either above or below the plane of the sugar (Figure 1.10B). In the C2´ -endo
Figure 1.10 Changes in nucleotide configuration that can affect the conformation of the double helix.
(A) The structures of anti- and syndeoxyadenosine. The two structures differ in the orientation of the base relative to the sugar component of the nucleotide, rotation around the β-N-glycosidic bond converting one form into the other. The three other nucleotides also have anti and syn conformations. (B) Sugar pucker, illustrating the positioning of the sugar carbons in the C2´ -endo and C3´ -endo
(A) Rotation around the β-N-glycosidic bond
(B) Sugar pucker
configuration the 2´-carbon is above the plane and the 3´-carbon slightly below, and in the C3´ -endo configuration the 3´-carbon is above the plane and the 2´-carbon below. Because the 3´-carbon participates in the phosphodiester bond with the adjacent nucleotide, the two pucker configurations have different effects on the conformation of the sugar–phosphate backbone.
Conformation changes resulting from rotation around the β-N-glycosidic bond and sugar pucker can give rise to major changes in the overall structure of the helix. It has been recognized since the 1950s that changes in the dimensions of the double helix occur when fibers containing DNA molecules are exposed to different relative humidities. For example, the modified version of the double helix called A-DNA has a diameter of 2.55 nm, a rise of 0.23 nm per base pair, and a pitch of 2.5 nm, corresponding to 11 base pairs per turn (Table 1.1). Like the B-form, A-DNA is a right-handed helix and the bases are in the anti-conformation relative to the sugar. The main difference lies with the sugar pucker, the sugars in the B-form being in the C2´ -endo configuration, and those in A-DNA in the C3´ -endo configuration. Other right-handed variations of the double helix include B´-, C-, C´-, C´´-, D-, E- and T-DNAs.
A more drastic reorganization is also possible, leading to the left-handed Z-DNA, in which the sugar–phosphate backbone adopts an irregular zigzag conformation. Z-DNA is a more tightly wound version of the double helix with 12 bp per turn and a diameter of only 1.84 nm ( Table 1.1). It is known to occur in regions of a double helix that contain repeats of the motif GC (i.e., the sequence of each strand is ..GCGCGCGC..). In these regions, each G nucleotide has the syn and C3´ -endo conformations, and each C has the anti and C2´ -endo conformations. The bare dimensions of the various forms of the double helix do not reveal what are probably the most significant differences between them. These relate not to diameter and pitch, but to the extent to which the internal regions of the helix are accessible from the surface of the structure. As shown in Figure 1.9A , the B-form of DNA does not have an entirely smooth surface: instead, two grooves spiral along the length of the helix. One of these grooves is relatively wide and deep and is called the major groove; the other is narrow and less deep and is called the minor groove. A-DNA also has two grooves (Figure 1.11), but with this conformation the major groove is even deeper, and the minor groove shallower compared with B-DNA. Z-DNA is different again, with the major groove virtually nonexistent but the minor groove very narrow and deep. In each form of DNA, part of the internal surface of at least one of the grooves is formed by chemical groups attached to the nucleotide bases. In Chapter 11, we will see that expression of the biological information contained within a genome is mediated by DNA-binding proteins that attach to the double helix and regulate the activity of the genes contained within it. To carry out their
TABLE 1.1 f EATURES O f THE DI ffERENT CON fORMATIONS O f THE DNA DOUBLE HELIX
Another random document with no related content on Scribd:
A Moment of Suspense
Perhaps I may mention that never in my experience have I seen an animal more terrified than that [35]pony was on that occasion. When we stood him again upon terra firma his legs quivered so that it was quite painful to witness, and he neighed and neighed convulsively, and made the sound echo all down the valley. I may also add that from that same scaffolding, that same morning, one of the workmen had fallen over, and his body was dashed into an unrecognisable mass on some rocks in the foaming stream below.
I had heard most peculiar rumours from the natives that the Tibetans had come in great force on to British territory at Garbyang, and with great pomp and much beating of drums were proclaiming that the British Government were afraid of coming to Tibet, and that the entire Bias and Chaudas had now been ceded to Tibet.
It may be remembered that as early as 1898, after the return from my first journey to Tibet, I pointed out to the Government the abuses of Tibetan officials who came over to our side of the frontier, claimed Sah-tal or Land Revenue from British subjects, and even tortured unprotected natives on our side of the frontier if they refused to pay. Some prominence was given in the House of Commons to questions on this subject, and the Governor of the North-West [36]Provinces had to acknowledge that such was the case, but declared that steps would be taken to stop the abuses. That the Sah-tal was paid was admitted, but it was stated to be a trade tax, which was untrue, and possibly uttered in ignorance of facts. It was agreed that the payment of the Sah-tal by British subjects should, nevertheless, be stopped, and it was actually suspended.
Apart from the fact that the words Sah-tal in themselves mean “land revenue,” and that it is paid on the amount of land possessed by Shokas on British soil, a further proof that it is a land tax is provided by the case of Darma Shokas, who pay the Sah-tal on their land to the Jong Pen of Taklakot, and who do no trading with that particular part of Tibet.
Furthermore, the Jong Pen of Taklakot receives the amount with the distinct understanding that it is a land tax, and not a trade tax, and as such
it appears in Government reports by the few British officials who had been to the frontier, such as Mr. Sturt, Mr. Larkin, and other deputy commissioners and collectors, and the Political Agent on the spot. In Darma the Sah-tal was collected direct by Jong Pen’s emissaries, but in Bias and Chaudas the Political Agent was made to act as a servant [37]to the Tibetans, and after collecting the revenue handed it over to the Jong Pen’s officials at Gungi and at Garbyang, two villages conveniently situated for the purpose.
Chaudas, being the most distant from the Tibet border, only paid a nominal sum of 11 rupees in cash, and some 14 rolls of rough cloth; but Garbyang, for instance (besides a real trade tax jointly with Chaudas of 370 boxes of grain, each containing some 24 lbs., and 14 boxes of coarse cloth), paid for Sah-tal the following items:—
86 (three anna bits).
Oats 1230 lbs.
Saluk (right of pasture) 2 Tibetan rupees.
Ghur (sweet paste) 4 balls.
Coarse cloth
Shoka liquor
Timber
3 entire rolls.
55 jars.
30 huge logs.
Iron 4 lbs.
Cakes, sattoo, charcoal, and liquor for collecting party.
The village of Kuti also paid a heavy Sah-tal.
Rupees 17
Oats 1210 lbs.
Saluk 2 rupees.
Garah cloth 40 yards.
Haunches of mutton 12
Jars of chökti (wine) 50
Ghur 28 balls.
Iron 2 lbs. and charcoal, cakes, meat, liquor, etc.
[38]
Darma paid some 150 balls of ghur, which had a local value of about 75 rupees.
Notwithstanding that the Government of the North-West Provinces could not or would not see matters in the right light, it was a great gratification to me to think that, on my giving publicity to such an injustice, the Government had at last deemed it right to discontinue the payment of the land revenue to Tibetans by our frontier subjects. It came, therefore, as a very great surprise to me, on approaching Garbyang, to hear that the Tibetans had come over to that place in great force,—with the sanction of the British Government,—and that several officials, including a Nerba, some Dhats in red and black tunics and long swords, four jimidars, two drummers, and a number of soldiers were parading the streets, proclaiming the annexation of the district to Tibet, and ill-using the natives who differed. They were gorging themselves at the expense of the natives, and having a fine time all round.
At the request of a deputation of Shokas, I hastened on to Garbyang; but the Tibetans, believing that I was coming up with a large and well-armed expedition to take a revenge upon them for former misdeeds, deemed it advisable to pack [39]up their chattels, pocket their pride, and decamp over the frontier.
I met with a most enthusiastic reception from the Garbyalis, who wished me to remain in their village until the passes were closed up by snow so as to keep their compulsory guests away. The Shokas seemed depressed and disgusted at what they rightly believed unwarrantable treachery on the part of the Government of India. They had always been faithful—even under trying circumstances—to the British, and such undeserved treatment led them to the one conclusion, preached by full-powered
Tibetan lungs, that England must indeed be weak, or at least afraid of Tibet.
Undoubtedly these sad and unchecked occurrences all along our Tibetan frontier—you see, it is so uncomfortable for Anglo-Indian officials to go up to the snows to make inquiries and so on—have been the chief cause of Tibetan aggressiveness. There is no man so brave as the one who believes his opponent is not going to fight; and perhaps now, after the much-trumpeted, enigmatical expedition to Lhassa and the unexpected sudden retreat from that city, they will be no better than before.
I was unable to accede to the request of the natives to remain to protect their village, as I [40]intended going farther; and, moreover, the Government of India having become alarmed at my proceeding a second time towards Tibet, behaved with a considerable amount of amusing childishness. Government chaprassis were arriving in quick succession with secret orders to prevent natives selling me provisions, to induce my men treacherously to abandon me, steal my weapons and food, and place every possible obstacle in my way
Some native friends, who, according to custom, had presented me with a ball of ghur and two handfuls of rice, were arrested and prosecuted, being brought down to Almora, some 145 miles, for the purpose. Others met with considerable annoyance, accusations of all kinds being showered upon innocent people—even such gross nonsense as accusations of hiding quick-firing guns and pieces of ordnance!
Both the Governor and the Commissioner at Naini-Tal, and the Deputy Commissioner of Almora, seemed quite hysterical over the affair, and some absurd threatening letters were written to me. My mails were invariably tampered with, and all letters opened. Among the most amusing requests may be noted one that I should show a Chinese passport if I wanted to traverse Bhot (British territory), [41]which made it seem as if the cession of that territory to Tibet had really been accomplished—although even then, as I unfortunately experienced before, a Chinese Imperial
passport has as much effect upon Tibetan officials as a piece of wastepaper would on the Prime Minister of Great Britain.
I was ordered to divulge my plans, which of course I refused to do, principally because I had no plans to divulge; and, further, was told I must stop and “wait for Government orders.” I replied that, being nobody’s servant, I would receive no one’s orders. I had not asked nor wished nor needed Government assistance, and I would go when and where I liked.
I was very sorry that the Political frontier Agent—a most sensible man— was subjected to much inconvenience and anxiety by constant orders to stop my expedition by any means—a matter in which I also happened to be concerned. He could not, as ordered, starve out my expedition, because I had plenty of food of my own. Silly as the orders given him were, he obediently did his utmost to check my movements, and, moreover, behaved like a gentleman into the bargain.
Some spies whom I found in my camp came in [42]for some severe punishment, and two Government chaprassis, who had one night been despatched to threaten my men and induce them to desert and capture my food and ammunition, received a sound thrashing, were disarmed, and I took them prisoners and brought them along with me to do some mountaineering! At the same time I sent word to those in authority that, when it should suit me, on my return, I would personally hand over the two prisoners.
Further orders arrived that any one selling or giving food or showing friendship to me should be arrested and sent down to Almora for punishment. All this seemed very useless, as I possessed all I needed; but, in order to prevent further persecution to some of my friends, I decided to cross the boundary at once—the Kali River—and enter the Nepal kingdom—a country also closed to foreigners, as everybody knows. [43]
CHAPTER V
In the meantime the Tibetans, who had hastily retreated across the boundary into their country, had given the alarm of the approach of my expedition. A spy I had sent over reported that great commotion reigned at Taklakot. Bridges had been hastily destroyed, stone barriers were put up, while warlike preparations of all kinds were carried on in feverish speed. A bluffing message was sent over by the Tibetans to inform the Political Agent that 5000 men were ready to meet my force and capture me again, when they would cut off my head and throw my body into the river—a threat which was by no means new to me.
This sounded interesting, only the Tibetans forgot that they would have to catch me first! The passes—both the Lippu and the Lumpiya (through the latter of which I had evaded their vigilance on my first journey)—were, they stated, [44]strongly guarded. I well knew the contemptible cowardice of the Tibetans, and I took but little notice of these threats; for this time— unlike my first journey—I had twenty men, all of my own selection, and they were magnificent fellows—faithful and plucky to quite an unusual degree.
Red Lamas
In answer to these Tibetan messages to the Government, it is possibly of some interest to notice that the Government of the North-West Provinces in all haste informed the authorities of Taklakot in words to this effect:—
“Mr. Landor is coming to Tibet. Possibly he will enter the country. The Government of India has taken steps to prevent any men accompanying him, also to take away his baggage, provisions, arms, and ammunition. He will enter alone and will be at your mercy. We [the Government] are in no way responsible for his entering Tibet.”
This message surprised me even less than the Tibetan effort. Indeed, I well knew that no one more than some officials of the North-West Provinces wished me dead. You see, I had on several occasions shown them up in a very poor light, and now again I was discovering fresh and most unaccountable misdeeds on their part all along the line. Naturally, I quite understood that it was an [45]uneasy conscience and the fear of being exposed which prompted them to act in such a childish manner in order to prevent my seeing what was going on.
The Political Agent, who was constantly receiving orders which he was powerless to carry out, such as arresting me,—which I defied him to do,— seemed in much distress, and to relieve the strain upon him I decided to cross over into forbidden Nepal territory The British authorities had taken the precaution to warn the Nepalese also that I might cross over their boundary, that my conduct was not countenanced by the Government, and that soldiers must be sent to stop me and possibly capture me. This led to some incidents which gave us a good deal of unexpected merriment.
A guard of Nepalese soldiers was hurried up, and, unaware that it was there, I crossed over the boundary, with our rifles in their cases for protection against the rain, and not at all prepared for war. Two of my men, who carried a rifle each, were a distance ahead. On nearing a small fort and guard-house they were pounced upon by Nepalese soldiers who were hiding behind some rocks, the rifles were seized, and the Nepalese escaped with them into the fort up higher upon the hill. [46]
My men ran back to me in a great state of excitement to report the occurrence. We hurriedly prepared a plan of attack, and without losing any time climbed up to the fort. Upon the walls some gaily attired figures peeped over, brandishing kukris and a few obsolete rifles. I could hear them behind strengthening the hastily barricaded door. An officer peeped over and asked what we wanted.
“I want my rifles back.”
Tibetan Man spinning Wool
“Your rifles are not here. We have already despatched them to Katmandu” (the capital of Nepal).
“Very good. If you do not return my rifles within two minutes we will come and get them ourselves.”
“We will fight you.”
“All right.”
Pounding with big rocks and by the aid of a wooden lever we got the door loosened, and while the garrison’s attention was directed to prevent our entering that way, four of my best men and I climbed over the wall on the opposite side of the fort and covered the defenders with our rifles. I demanded that they should lay down their weapons or I would shoot. After some indecision, most of them did. The others, especially one who fired at [47]us, got a severe pounding and were disarmed by my men, who had now all entered the fort.
We ransacked the place, and eventually recovered the captured rifles, after which we bade the Nepalese a respectful salaam and proceeded on our way.
“But,” humbly put in the trembling Nepalese, “you cannot travel on Nepal territory. It is forbidden to foreigners, and I have orders to stop you.”
“Very well, do it!”
“I cannot,” he meekly mumbled, as he rubbed aching bruises he had received in the encounter. “But,” he said, “I will follow your movements.”
“Oh, you can follow any movements you like, but mind you do not come too near.”
“Oh, no, no, sahib!” he exclaimed.
The guard, in fact, followed us for some days—and always at a most respectful distance—until we got high up among the snows and glaciers.
Then we lost sight of them. Anyhow, I had no further annoyance all through my journey in Nepal, and found the few natives we saw quite attractive, picturesque, and kind.
After leaving the fort we travelled practically south-east over very rough country, my objective [48]being to visit and if possible climb one of the Lumpa peaks which towers in a majestic needle above most other mountains in the neighbourhood. Every now and then, when we got to some higher point of vantage, we got a beautiful view of it. My men—to my astonishment—received my plan with enthusiasm, for they seemed to hold the snows in veneration. Little they then knew how much they would have to suffer upon them.
In that portion of North-West Nepal the population is sparse and somewhat mixed. Perhaps the finest and most interesting types I saw were the cross-breeds of Shokas and Nepalese, which seemed to produce most striking heads. One of the coloured drawings representing a typical Nepalese-Shoka young man, with temples shaved and long wavy hair hanging down his back, is, I think, a good example. These men had very fine traits about them, were most hospitable and civil, manly and serious in manner. They were enterprising traders, carrying on a brisk business mainly in borax, salt, wool, and skins, which they generally bartered with Tibetans in exchange for grain, ghur, and other food stuffs.
A Nepalese Shoka
It is rather interesting to note that whereas similar frontier tribes on neighbouring British soil [49]are imposed upon, taxed and ill-used by Tibetans, no interference at all is experienced by those living under the protection of the King of Nepal. Possibly this is due to the fact that when Nepalese subjects have been ill-used in Tibet an armed expedition has been sent over the frontier by the Nepalese, and reparation demanded and obtained at once.
Nepalese women are not unattractive, having large brown eyes, made additionally languid by blackening the lower lid, long eyelashes and wellcut features, skin of a well-polished, smooth, light-yellowish brown, and most graceful hands and feet. The better class are generally much decorated with heavy silver necklaces, bracelets, and rings, and with huge nose-rings of gold, brass, or silver. They are handsome while young, but they fade away at an early age, and the smooth skin becomes wrinkled and grooved long before they are thirty. They are said to be most affectionate, and of a somewhat jealous temperament, which they couple with extreme conjugal fidelity.
In Western Nepal the hair is worn in many little plaits festooned on both sides of the forehead, and a head-cover, white, yellow, or red, such as is seen in India among Hindoo women, is usually worn.
The women have comparative freedom, and [50]seldom are they seen completely veiled. Sati or suttee, the widow’s self-sacrifice by throwing herself upon the flames which have cremated her dead husband, is now forbidden by law in Nepal. It is, nevertheless, indulged in to some extent in districts far away from the capital.
Nepal is principally known to English people as the country from which, by a special agreement, we draw recruits for our Gurkha regiments. It should, nevertheless, be borne in mind that not all people living in Nepal are by any means Gurkhas—far from it. We find in Nepal a great variety of types —some pure enough, others distinct mixtures of two or more races.
The Gurkha district proper lies in the north-easterly portion of the Gandak basin, the chief city being Gurkha, some fifty-four or fifty-five miles to the west of the Nepalese capital, Katmandu.
Formerly Nepal’s sovereignty extended over Kumaon, which we have already traversed, as far as the Sutlej River. The Nepal kingdom, as it stands now, is an elongated tract of country—mostly mountainous—some 500 miles in length, and less than 150 miles in breadth at its widest points. As everybody knows, it is situated on the southern slopes of the central Himahlyan range, [51]and borders with Tibet on the north, Kumaon (NorthWest Provinces) to the west, the Kali River, as we know, forming the boundary; the North-West Province and Bengal to the south, and Sikkim to the east.
Nepal is divided into three natural zones by high ranges with peaks rising from 25,000 to 28,000 feet, these ridges shooting off southwards from the main Himahlyan range. The eastern zone is drained by the Kosi River, the central by the Gandak, and the Western by the Gogra. To the above should be added what is called the Terai and the thickly-populated Nepal Valley, which, geographically, must be classed separately. Formerly Nepal was divided into twenty-four principalities in addition to the Gurkha kingdom, but since the invasion by the latter of the entire country they have been divided into five provinces—viz. Gurkha, Malibam, Palpa, Pokra, and Khachi.
In the space at my disposal I cannot enter into the history nor go into a detailed description of the many tribes which inhabit Nepal, but generally speaking, in the aboriginal stock a Mongolian origin is evident, particularly in the population of the central zone, where recent Tibetan influence is marked; but this is not the case with the [52]inhabitants of the lower region, who seem to have descended partly from a mixture of an earlier Tibetan and Aryan source. Perhaps among the most prominent aboriginal tribes we should mention the Magars, Gurungs, Newars, Sunwars, the Khambas or Khambus, and Yakhas (the latter two closely resembling Rais), the Yakthumbas, Murmis, and Lepchas. Then in the forests of Western Nepal we find to this day wild tribes of Chepangs and Kasundas.
In the Terai district live the Tarus, a servile and ill-shaped people of no great strength of character; in the eastern zone we have Limbus, a name
A Nepalese Lady
which practically includes Kirantis, Ekas, and Rais—a people of apparently Tibetan origin, flat-faced, beardless, and with long unplaited hair, who wear wide trousers and a short jacket instead of the long coat of the Lepchas.
The central zone, from which most of the recruits for the British Gurkha regiments are drawn, is inhabited by a small but strongly built, warlike, and plucky people, the Magars and Gurungs, the Magars being in their turn subdivided into six distinct tribes, of which the Thapa tribe is the most numerous. Both the Magars and the Gurungs—who are somewhat taller and more [53]stoutly built than the Magars—although of marked Mongolian characteristics, are by religion Brahmins, but have no sort of strong caste prejudices such as one finds, for instance, in India. Gay and simpleminded, with hearts of gold, faithful and obstinate to an extreme degree, independent in themselves yet loyal to their friends or employers, fond of fighting and possessing the courage of lions, these fellows make wonderful soldiers indeed. Perhaps in many ways they are not unlike the Japanese, both in appearance and temperament, except that the physique of the Gurkha is the superior of the two.
In the Valley of Nepal—or Nepal proper—we find mostly Newars and Murmis. The Newars, who claim to be the aborigines of the Nepal Valley, are in many ways not unlike the Bhotias or Shokas, and in many of their customs and in their habitations show a marked similarity to those tribes of Mongolian descent. Their language resembles Tibetan, and their religion is a modified form of inherited Buddhism. The Newars are much given to agricultural pursuits, and they do not display so prominently the fighting qualities of the Magars and Gurungs. On the contrary, they seem to devote all their energy to art and trade, of which [54]they seem to have the entire monopoly in Nepal. The Murmis resemble Bhotias even more closely than the Newars. They are subdivided into two classes—the Barathamang, which is the most numerous, and the Atharajat.
Katmandu, the capital of Nepal and residence of the King and Government, lies in this valley, and is a city of great size and some beauty
The Terai is inhabited by a poor, sickly-looking race called the Tarus, malarial fever being rampant in their district, while wild beasts abound in the jungle. They seem to be a poverty-stricken lot with a wretched physique—although endowed with astounding strength and powers of endurance. They divide their time between fishing, agriculture, and hunting wild animals.
Now comes the western zone, which is entirely inhabited by non-Gurkha tribes, such as the Doti and Jumli (the Doti’s country being south of the Jumli’s), as well as by wilder tribes, such as the Chepangs and Kusundas, the former being very similar to the Raots or Rajis of Kumaon. Then there are minor Hinduised tribes generically known as Parbatiyas (or hill men).





















































































