biology
Review


Review
GianlucaPolgar*,MattiaIaia,TommasoRighiandPietroVolta
WaterResearchInstitute(IRSA)—CNR,LargoTonolli50,28922VerbaniaPallanza,Italy; mattia.iaia@irsa.cnr.it(M.I.);tommaso.righi@irsa.cnr.it(T.R.);pietro.volta@cnr.it(P.V.)
* Correspondence:gianluca.polgar@irsa.cnr.itorgianluca.polgar@gmail.com
SimpleSummary: Inagreatpartoftheworld,troutfishinghaslonginspiredhumanspiritual idealsofimmersionintonatureandrecreation,farremovedfromthefast-encroachingurbanization. Concurrently,thesevaluesandemotionsfueledawhite-hotbusiness,establishingafloridmarket ofoutdoorrecreation.Sincethe20thcentury,thetrout-cultureindustrystrivedtoprovideanglers withfishingentertainmentbystockingmassiveamountsofnon-nativetroutsindozensofcountries, irrespectiveofthelakes’andrivers’carryingcapacity.Thishaddireconsequencesonthestructural andfunctionaldiversityoftheseecosystems.“Troutwars”sparkedthroughouttheworldbetween thepromotersofstockingactivitiesandthepromotersof“wildtroutmanagement”andethics. The“Italiantroutwar”hasbeenfoughtontheharshbattlegroundoftrouttaxonomy,ecology, distribution,andnativevs.non-nativeinterfertilespecies.NorthernItaly,hometotheItalianAlpine andsubalpinetroutsandeconomiccenterofthenationaltrout-cultureandstockingindustry,was particularlyaffectedbythisclash.Wereviewherethestateofartofthisongoingdebate,outliningour scientificviewofthetaxonomy,evolution,distribution,andsustainablemanagementofthenative ItaliantroutsofnorthernItaly.
Citation: Polgar,G.;Iaia,M.;Righi, T.;Volta,P.TheItalianAlpineand Subalpinetrouts:Taxonomy, Evolution,andConservation. Biology 2022, 11,576. https://doi.org/ 10.3390/biology11040576
Received:21February2022
Accepted:8April2022
Published:11April2022
Publisher’sNote: MDPIstaysneutral withregardtojurisdictionalclaimsin publishedmapsandinstitutionalaffiliations.

Copyright: ©2022bytheauthors. LicenseeMDPI,Basel,Switzerland. Thisarticleisanopenaccessarticle distributedunderthetermsand conditionsoftheCreativeCommons Attribution(CCBY)license(https:// creativecommons.org/licenses/by/ 4.0/).
Abstract: Duringthelast150years,thetrout-cultureindustryfocusedonenhancingtroutpopulations bystocking,inresponsetothegrowinganglers’demandandthehabitatdegradationassociatedto therapidurbanizationandhydropowerdevelopment.TheindustrializednorthofItaly,hometothe ItalianAlpineandsubalpinetroutpopulations,isthesourceofmostoftherevenuesofthenational trout-cultureindustry.Itsrapidgrowth,andthemassiveintroductionofnon-nativeinterfertiletrouts erodedthegeneticdiversityofnativelineages,leadingtoharshconfrontationsbetweenscientists, institutions,andsportfishingassociations.Wereviewherethestateoftheartofthetaxonomyand distributionofthenorthernItaliannativetrouts,presentingbothscientificresultsandhistorical documentation.Wethinktheonlynativetroutsinthisregionare Salmomarmoratus,widespreadin thisregion,plussmallandfragmentedpopulationsof S.ghigii,presentonlyintheSouth-western Alps.Westronglyrecommendtheinterruptionofstockingofdomesticatedinterfertilenon-native troutsinthisarea,andrecommendtheadoptionofEvolutionarySignificantUnitsforsalmonid fisherymanagement.Wefurtherproposefutureresearchdirectionsforasustainableapproachtothe conservationandecosystemmanagementofthefisheryresourcesandinlandwatersofnorthernItaly.
Keywords: recreationalfisheriesfisherymanagement;introgressivehybridization;stocking; non-nativespecies;allochthonousspecies;troutfishing;trouttaxonomy
1.Introduction:TheGlobalCulturalValueofTroutandthe“TroutWars”
InEurope,freshwateranglingoriginatedassubsistenceandsmall-scaleartisanal fisheriesservinglocalmarkets,growingalongsidesportfishingasearlyasintheLate MiddleAges,virtuallybecomingasynonymoftroutfishing[1–3].Analogoussubsistence trout-fishingculturesalsodevelopedinhuntingandgatherersocieties,suchastheNative Americancultures[4].
Sportfishingandtroutflyfishinggreatlyexpandedduringthe19thcentury,spreading throughouttheworldthroughEuropeancolonialism(America,Asia,Africa,Australia, andNewZealand)andtheriseofthenation-states,associatingwithcultural,economic, spiritual,andrecreationalvalues[3].Artificialpropagationwasscientificallydescribed andpopularizedinFranceinthe1840s–1850s.IntheEuropeanimperialisticsocieties, highlyinterconnectedbysteamshipsandrailroads,thesepropagationmethodsfueleda burgeoningoutdoorrecreationindustry,i.e.,theconsumptionofnatureforleisure-class consumers[5].Thissportingculturegaverisetopresent-dayfisherymanagementand angling,makingtroutaglobalizedcommoditythroughtheintroduction,acclimatization, andnaturalizationofnon-nativetroutspeciesprovidedbyprivateandgovernmental associations.TheEuropeandiasporatoothercontinentsfollowingWorldWarIfurther acceleratedthisprocess.“Salmonizing”becameasynonymofacclimatization[6].The firstItalianhatcherywasestablishedinPiedmontin1859[7],andexperimentsofartificial propagationandacclimatizationofnon-nativefishes,namely Coregonus and Salvelinus speciesfromSwitzerlandandGermany,wereconductedinseveralsubalpinelakesduring the1860s–1890s[8].Introductionsandtranslocationsacceleratedwiththeestablishmentof thetwoItalianfisherycentersinBresciaandRome,in1893–1895,withjurisdictionover thePadano-VenetianichthyographicdistrictandtherestofpeninsularplusinsularItaly, respectively[9,10].Massiveacclimatizationandtranslocationactivities(~12billionfishes ofdifferentspecies,including16non-nativespecies)wererapidlyimplementedinthe following50years,slowingdownafter1945[10].
Duringthe20thcentury,thepoliticalimpetustomeetthegrowinganglers’demand formorefishbyturningrecreationalfishingintoaprofitableeconomicsectorfurtherpromotedthefish-cultureindustry,whichincreasinglyfocusedonenhancingbothnativeand non-nativefishpopulationsbystocking.Thesynergisticeffectsofmassivestockingand thehabitatdegradationassociatedwiththeongoingurbanizationincreasinglyimpacted freshwatersystems,eventuallytriggeringdramaticconservationandmanagementissues. Artificiallystockingmoremanufacturedfishthantheecosystems’carryingcapacityand redefiningnatureasaplaygardentemporarilyallowedtoignoretheongoingdamagetothe ecosystems[3,11].Inparticular,massivestockingofnon-nativetroutsstartedtoseverely impactnativeassemblages,bothecologicallyandgenetically,duetothewidespreadinterfertilityamongsalmonidtaxa,e.g.,[12].
Inthe1960s,afterWorldWarII,themodernecologicalmovementandtheconceptsof wildtroutmanagementandethicsgraduallydevelopedintheU.S.A.alongsideput-andtakefisherypractices,initiallyfocusingonpollution,habitatdegradationandfragmentation,and,then,extendingtofishingregulationsandrestorationofwildnativepopulations. Harshconfrontationsbetweenconservationistsandadvocatesofstockingsparkedeverywhere,fromU.S.A.toSouthAfrica,beingdescribedasthe“troutwars”[3].
InItaly,stockingactivitiesdramaticallyincreasedinthe1970s,afterthemanagement ofinlandwaterspassedtotheProvincesin1974[9,10].Foratleastthesubsequent25years, stockingwasconductedbyfewlargeaquaculturefacilities.Thelargestone,locatedin theVenetoRegion,waswidelyusedbyseveralnorthernandcentralItalianProvincesto stockmassiveamountsofseveralpoorly-determinedfishspeciesfromthePadano-Venetian ichthyographicprovince,vaguelydefined“pescebianco”(literally:“whitefish”[9]).More recently,governmentcompliancewithEuropeanlegislation[13]confirmedtheprohibition tostocknon-nativefishspeciesandpopulationsintoItalianwaters,unlessasite-specific environmentalimpactassessmentdemonstratesthelackofnegativeimpactsonnative speciesandtheenvironment[14,15].ThegovernmentproposedareferencelistofItalian nativeandnon-nativespeciesofinterestinthefisheriessector.Amongthenon-native fishesthereisoneoftheworld’sworstinvasivespecies,theAtlanticbrowntrout S.trutta Linnaeus1758[16],whichhasbeenthebackboneofthestockingandsportfishingItalian industriesforalmosttwocenturies.Asaconsequenceofthestrongsocialandpolitical tension,arecentamendmenttothenationalBudgetlaw[17]hasbeenapproved[18]to suspendtheapplicationoftheselaws[14,15]until2023.Aharshconflictistakingplace
betweenstakeholdersprioritizingthegeneticandecologicaldiversityofnativepopulations, mainlyincludingfisheryscientistsplussomeanglersandmanagers,andthoseprioritizing theexploitationofinlandfisheries,mainlyincludingfishculturist,anglers,sportfishing associations,thehydropowerprivatesector,andthemajorityoftheProvinceandRegion administrations.ThisclashwasparticularlydramaticintheindustrializednorthofItaly, hometotheItalianAlpineandsubalpinetroutpopulations,andsourceofmostofthe revenuesofthenationalstockingandanglingindustries.This“Italiantroutwar”also provokedheateddebatesonthetaxonomicandgeneticidentityofthestockedtrouts, e.g.,[19],andonthenativestatusoftroutspeciesinItaly,e.g.,[20–23].Since5–10years, severalItalianfish-culturecompaniesstartedtointroducemassiveamountsofnon-native stocksofpeninsulartrout(Section 2)inthisandothergeographicareas.Suchstocksare collectedfromseverallocationsinpeninsularandinsularItaly,includingdomesticated progenyoftenhybridizedwithnon-nativeAtlanticstocks[19].Thesearebeingsold as“Mediterraneantrout”,e.g.,[24],andcurrentlyadvertisedasaconservation-friendly alternativetothenon-nativeAtlanticbrowntrout.Thiscommercehasalsobeenfacilitated bytheidea,arguablyunsupportedbyanyscientificevidence,ofawidespreadpresence ofviablepopulationsofapeninsulartroutlineageinnorthernItaly(i.e., S.cenerinus; Sections 2 and 4).
Ontheotherhand,thecurrentglobalinterestintrouts,facilitatedbysocialplatforms andInternetvirtualcommunities,initiatedseveralenvironmentalethicsandangler-driven conservationinitiatives(e.g.,TroutUnlimitedintheU.S.A.;BalkanTroutRestorationGroup inSlovenia;numerousEuropeanLIFEprojects).Sharedattachmenttoplace,characterizing awiderangeoffishenthusiasts,fromanglerstofisheryscientists[3],couldraisescientific awarenessandfostercollaborationamongdifferentstakeholders.Thiscoulddevelopglobal networksofmultiplepartiesaddressingissuessuchassustainability,wildmanagement, andtransboundaryconservationissues,suchasclimatechange,pollution,andobstaclesto fishmovements(e.g.,damsandhydroelectricpowerplants).
WereviewherethestateofartofthetaxonomyandevolutionoftheItaliantrouts, clarifyingourscientificviewofthesetopics,andproposingfuturedirectionsforascientific approachtotheconservationandecosystemmanagementofthefisheryresourcesand inlandwatersofnorthernItaly.
2.NativeItalianTroutsandtheTaxonomyofthe“PeninsularTrout”
Amongthevalidnominaltaxa[25]ofthenativetroutsdescribedintheItalianpeninsulaandthemajorItalianislands, Salmocettii Rafinesque-Schmaltz1810wasdescribed fromSicily(typelocality:ValDemoneinnortheasternSicilyandValdiNotoinsoutheasternSicily,notypesknown[25,26]). S.marmoratus (Cuvier1829)isasubendemism ofnorthernItalydescribedfromthe“lacsdeLombardie”(syntypesnotavailable[25,27]; Section 3). S.cenerinus Nardo1847wasdescribedfromnortheasternItaly(typelocality: notfarfromthesea,inriversdrainingtotheVenetianlagoon;notypesknown[25]).The originaldescriptionof S.cenerinus waswrittenfromthelate1700stotheearly1800sby S.Chiereghin,andpublishedposthumously[28];asummaryofthisdescriptionwasfirst publishedbyNardo[29]. S.macrostigma (Duméril1858)hasbeenconsideredbyseveral authorsasanItaliantrout;however,itwasdescribedfromNorthAfrica(typelocality: Oued-el-Abaïch,Kabylie,Algeria[25,30]). S.ghigii Pomini1941wasdescribedfromcentral Italy(typelocality:SagittarioRiver;notypesknown[25,31]). S.fibreni Zerunianand Gandolfi1990,describedfromtheLakePostaFibrenoincentralItaly,and S.carpio Linnaeus 1758,describedfromLakeGarda,arerestrictedendemismsdefinedbyecomorphological andgenetictraits[32].TheislandofSardiniamighthostanundescribed Salmo species[12]. Theshortdescriptionandillustrationof S.cenerinus [29]maycorrespondtothe pelagicmorphofseveralanadromous Salmo species([33,34],pers.obs.).Nardo[35] modifiedhispreviousview[29],raisingdoubtsontheoriginaldescriptionof S.cenerinus, acceptingtheviewof[36](citedas1858by[35]),andeventuallyconsideringthistaxon asasynonymof Truttafario L.(= S.trutta).HeckelandKner[36]reportedonlytwotrout
speciesfromtheVenetianProvinces: SalarAusonii Valenciennes1848[37](= Truttafario L.sensu[35])and Fariocarpio (= Truttacarpio sensu[35]= S.carpio)fromtheGardaLake. While Salargenivittatus HeckerandKner1858wassubsequentlyrecognizedasamorph of S.marmoratus [38],HeckelandKner[36]considered S.marmoratus asacolormorphof SalarAusonii.Therefore,Nardo[35]likelyconsideredthemarbletroutofthisareaascolor morphsof Truttafario.
Kottelat[38]assigned S.cettii tothenativepeninsularTyrrhenianandsouthernItalian trout,includingislands,and“tentatively”assigned S.cenerinus tothenativenorth-Italian (Adriatic)peninsulartrout.Consistently,hedidnotconsider S.cenerinus asjun.syn.of S. marmoratus,since“therewouldbenoavailablenameforthepresentspeciesanditshould beeitherlistedas Salmo sp.oranewnameshouldbecreatedforit”.Kottelat[38]also synonymized S.ghigii with S.cettii apparentlyonlybecausePomini[31]wasunableto discriminatethetroutsoftheSagittarioRiverfromtheSardiniantrouts.KottelatandFreyhof[39]acceptedthepointofviewofKottelat[38],whilenotingthat“recent studies(...) suggestthatthetroutsofSicily(... )belongtoadistinctmolecularlineage(... ).Ifconfirmed,thislineageshouldretainthename S.cettii;thename S.ghigii wouldprobablybe thevalidnameoftheothers”.
Usingcombinedmitochondrial(mtDNA)andnuclear(nDNA)markers,Segherloo etal.[12]foundacloserelationshipbetweentheSiciliantroutofValdiNotoandAtlantic S. trutta.AconsistentresultwasfoundbyanothernuclearphylogeneticstudyoftheMoroccan trouts,whichincludedtheSiciliantroutofValdiNotoinarobust“Afro-Atlanticclade”, likelyoriginatedfromacolonizationwaveofanAtlanticlineagefromIberia(“Duero” lineage;[40]).TheonlyNorth-AfricansampleanalyzedbySegherlooetal.[12],that theseauthorstentativelyassignedto Salmopellegrini Werner1931,iscloselyrelatedto MediterraneanandAdriaticbrowntrouts,thusclearlybelongingtoadifferentlineage;this samplecomesfromtheOumer-RbiaRiver,whereSnojetal.[40]identifiedtroutsofan “Atlasclade”.SeveralstudiesshowedthattheSiciliantroutismorphologicallydistinct fromotherItaliantrouts[30,41–44].MitochondrialphylogeniesalsoshowthattheSicilian troutisincludedinacladeofNorth-Africantrouts,whichalsoincludesthesequenced typesof S.macrostigma [45]andtheAtlantictroutlineage,calledthe“SouthernAtlantic clade”[22,46].However,nonuclearorcombinedmitochondrialandnuclearphylogenies wereeverconstructedincludingthetypesof S.macrostigma,whichmaybeunrelatedtothe Afro-Atlanticclade.
Rafinesque-Schmaltz[26]described S.cettii fromtwotroutpopulations:ValdiNoto andValDemone.Themolecularphylogenyof S.cettii hasbeeninvestigatedonlyusingthe formerpopulation,sincenogeneticsampleshaveeverbeencollectedandanalyzedfrom theValDemone,whichhaslikelybeenextirpated[47].Therecoveryandanalysisofany suchmolecularsample(e.g.,fromamuseumlot)wouldhaveimportantconsequenceson thescientificnamesofItaliantroutlineages.Therearethreepossiblescenarios:(i)theVal Demonepopulationbelongstoanundescribedendemictroutlineage;(ii)theValDemone populationisconspecificwiththepeninsulartrout;(iii)theValDemoneandValdiNoto populationsareconspecific(Figure 1).
Inthefirstscenario(Figure 1a),theValdiNotoandtheNorth-AfricanAfro-Atlantic clade(AAC,sensu[40])wouldbeclassifiedas S.cettii (olderthananyNorth-Africantrout taxonsofardescribed[48]),thepeninsularpopulationsas S.ghigii,andtheValDemone populationwouldbelongtoaundescribedspeciesthatwouldrequireformaldescription. Alsointhesecondscenario(Figure 1b),thecollectionofRafinesque-Schmaltz[26]contained twodistincttaxa,withtheValDemonepopulationbeingconspecificwiththepeninsular trout.Intheabsenceoftypematerial,itisarbitrarywhethertoassigneitherofthetwo sampledpopulationsto S.cettii.Therefore,therearetwopossibilities:either(a)Valdi NotoplusAACpopulationscouldbeclassifiedas S.cettii,andValDemonepluspeninsular populationsas S.ghigii,or(b)ValDemonepluspeninsularpopulationscouldbeclassified as S.cettii,andValdiNotoplusAACasoneofthefourNorth-African Salmo species (including S.macrostigma [48]),orasanewandyetundescribedspecies.Inthethird
scenario(Figure 1c),theSiciliantroutplusAACwouldbe S.cettii,andthepeninsular populationswouldbe S.ghigii.IntheabsenceofmaterialfromtheValDemone,weadopta classificationconsistentwiththethirdandmostparsimoniousscenario.
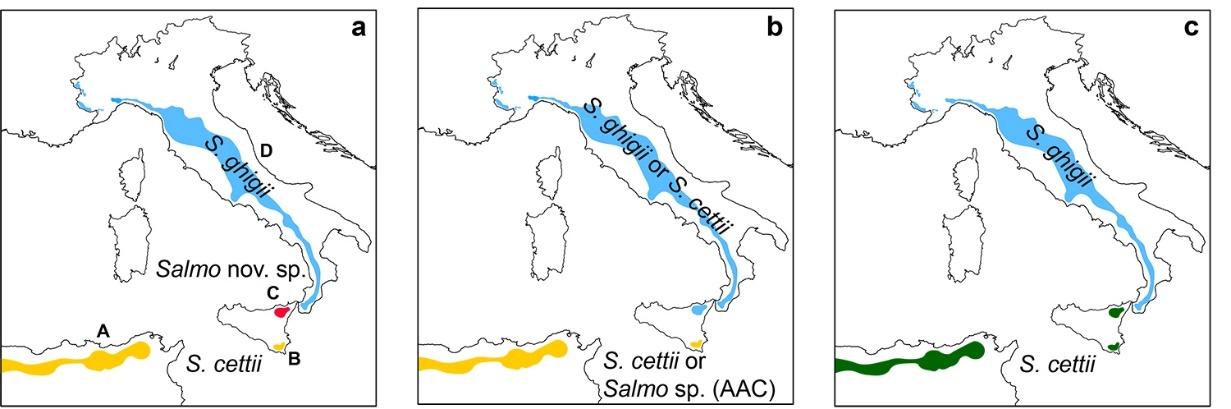
Figure 1. Three possible taxonomic scenarios (peninsular and Sicilian trouts, plus a North-African trout) if native trout samples are collected and analyzed from the Val Demone (no material presently available); (a) the native Val Demone population belongs to an undescribed and endemic lineage; (b) the native Val Demone population is conspecific with the peninsular trout; (c) the Val Demone and Val di Noto populations are conspecific. A: hypothetical distribution of a trout taxon of the “Afro-Atlantic clade” (AAC), conspecific with the Val di Noto trout population [40]; B and C: Val di Noto and Val Demone (likely extirpated [47]) populations, respectively, both described as S. cettii by Rafinesque-Schmalz [26]; D: peninsular trout, distribution range modified from [48]. No types are known for both S. ghigii and S. cettii. The taxon S. cettii is older than all North-African trout taxa [48].
Figure1. Threepossibletaxonomicscenarios(peninsularandSiciliantrouts,plusaNorth-African trout)ifnativetroutsamplesarecollectedandanalyzedfromtheValDemone(nomaterialpresently available);(a)thenativeValDemonepopulationbelongstoanundescribedandendemiclineage; (b)thenativeValDemonepopulationisconspecificwiththepeninsulartrout;(c)theValDemone andValdiNotopopulationsareconspecific.A:hypotheticaldistributionofatrouttaxonofthe “Afro-Atlanticclade”(AAC),conspecificwiththeValdiNototroutpopulation[40];BandC:Valdi NotoandValDemone(likelyextirpated[47])populations,respectively,bothdescribedas S.cettii by Rafinesque-Schmalz[26];D:peninsulartrout,distributionrangemodifiedfrom[48].Notypesare knownforboth S.ghigii and S.cettii.Thetaxon S.cettii isolderthanallNorth-Africantrouttaxa[48].
Withthelimitationofsubstantialsamplebiases,severalstudiesdidnotfindgeneticor ecologicaldiscontinuitiesbetweennativenorthern(South-westernAlps[49])andcentralsouthernpeninsulartroutlineagesthatwouldjustifythedesignationofdifferenttaxa, except S.carpio and S.fibreni [20,32,50–54].Segherlooetal.[12]assignedtroutsamplesofthe upperreachesofthePodrainageto S. cf. cenerinus,andsamplesoftheZrmanjaandMornos basins(Balkanpeninsula)to Salmofarioides Karaman1938,inthesameareaofitstype locality(KrkaRiver,Croatia[55];neotypedesignatedbyBianco[56]). S.cenerinus wasfound inbrackishconditions[28,29],howevertheonlynativeItaliantroutrecordedintheseais S.marmoratus [57].Further,therepresentlyarenoknownnativepopulations(norgenetic tracesofpastpopulations)ofpeninsulartroutintheareawhere S.cenerinus wasdescribed. Ontheotherhand,anadromousnon-nativepopulationsof S.trutta,includinghybrids, areknowntooccurintheAdriaticregion,includingItalianwaters[58,59].Chiereghin diedin1820,andlikelydescribed S.cenerinus fromthelate1700stotheearly1800s.Fish cultureprojects,possiblyincludingnon-native S.trutta,startedinthisareainthesecond halfofthe19thcentury[35].Ontheotherhand,thehypothesisthatthetroutdescribed byChiereghinwasapelagicmorphof S.trutta cannotberuledout,sinceintroductions ofnon-nativesalmonids,possiblyincluding S.trutta,haveoccurredinnorthernItaly atearliertimes(Section 4).Inthe1970s,BorroniandGrimaldi[8]justreportedthat introductionsofnon-native S.trutta hadbeenoccurring“fordecades”inItaly.Biancoand Delmastro[60]andBianco[56]synonymized S.marmoratus and S.cenerinus basedonthe illustrationof S.cenerinus [28],itsanadromoushabits[28,29],andinformationgleaned fromGridelli[61],whoreportedonlythepresenceofthemarbletroutintheVenezia GiuliaRegion,previoustostockingactivitiesofnon-nativebrowntrouts,whichstartedin 1934.However,Nardo’s[29]VenetianProvincesofthe1850s(typelocalityof S.cenerinus) aregeographicallydistinctfromGridelli’s[61]VeneziaGiuliaRegionofthe1930s[62]. Bianco[56]synonymized S.ghigii with S.farioides,however:(i)noneotypeof S.ghigii wasdesignated,likelyduetothedifficultyoffinding“purebred”individualsinthetype locality;(ii)nomolecularanalyseswereconducted;(iii)thesynonymywasessentially
In the first scenario (Figure 1a), the Val di Noto and the North-African Afro-Atlantic clade (AAC, sensu [40]) would be classified as S. cettii (older than any North-African trout taxon so far described [48]), the peninsular populations as S. ghigii, and the Val Demone population would belong to a undescribed species that would require formal description. Also in the second scenario (Figure 1b), the collection of Rafinesque-Schmaltz [26] contained two distinct taxa, with the Val Demone population being conspecific with the peninsular trout. In the absence of type material, it is arbitrary whether to assign either of the two sampled populations to S. cettii. Therefore, there are two possibilities: either (a) Val di Noto plus AAC populations could be classified as S. cettii, and Val Demone plus peninsular populations as S. ghigii, or (b) Val Demone plus peninsular populations could be classified as S. cettii, and Val di Noto plus AAC as one of the four North-African Salmo species (including S. macrostigma [48]), or as a new and yet undescribed species. In the third scenario (Figure 1c), the Sicilian trout plus AAC would be S. cettii, and the peninsular populations would be S. ghigii. In the absence of material from the Val Demone, we adopt a classification consistent with the third and most parsimonious scenario.
With the limitation of substantial sample biases, several studies did not find genetic or ecological discontinuities between native northern (South-western Alps [49]) and central-southern peninsular trout lineages that would justify the designation of different taxa, except S. carpio and S. fibreni [20,32,50–54]. Segherloo et al. [12] assigned trout samples of the upper reaches of the Po drainage to S. cf. cenerinus, and samples of the Zrmanja and Mornos basins (Balkan peninsula) to Salmo farioides Karaman 1938, in the same area of its type locality (Krka River, Croatia [55]; neotype designated by Bianco [56]). S. cenerinus was found in brackish conditions [28,29], however the only native Italian trout recorded in the sea is S. marmoratus [57]. Further, there presently are no known native populations (nor genetic traces of past populations) of peninsular trout in the area where S. cenerinus was described. On the other hand, anadromous non-native populations of S. trutta, including hybrids, are known to occur in the Adriatic region, including Italian waters [58,59]. Chiereghin died in 1820, and likely described S. cenerinus from the late 1700s to the early 1800s. Fish culture projects, possibly including non-native S. trutta, started in this area in the second half of the 19th century [35]. On the other hand, the hypothesis that the trout described by Chiereghin was a pelagic morph of S. trutta cannot be ruled out,
basedoncolorationpatternsandbiogeographicalreconstructions.Therefore,wechoose nottoconsider S.ghigii asajuniorsynonymof S.farioides.Asaresult,untilevidence isprovidedofintroductionsofnon-native S.trutta intheareawhere S.cenerinus was described,weconsider S.cenerinus asjun.syn.of S.marmoratus sensuBianco[56].While geneticdifferencesbetweenpeninsular-troutpopulationshavebeenfoundatdifferent geographicscales[23,63],untilmorecomprehensivegeneticandecologicaldataaremade availableonTyrrhenianandAdriaticnativeItaliantrouts,wetentativelyconsider S.ghigii asavalidnameforallthepopulationsofItalianpeninsulartrout,sensuZanettietal.[64] andLorenzonietal.[65].
Nuclearphylogeneticreconstructionsandmolecularclocksdefinedarobust S.marmoratus clade,includingtwodistinctnorthernandsouthernAdriaticcladesthatdiverged ~0.84 ± 0.4millionyearsago(mya)[66,67],andwhosetaxonomicstatushasnotyet beenevaluated.Pustovrhetal.[67]showedthat S.marmoratus iscloselyrelatedtoanuclear “S.trutta complex”lineage,includingseveralcladesassociatedwithdifferentbrowntrouttaxa,andestimatedthedivergencebetweenthesetwolineagesat 1.4 ± 0.8mya (2.2–0.6mya).Afossil-calibratednDNAphylogenyestimatedanearlierdivergence,at ~4–5mya[68].AnextensivemolecularphylogenycombiningnDNAandmtDNAsequencesrootedwith S.salar,essentiallyconsistentwithpreviousnDNAphylogenetic reconstructions,supported S.marmoratus asaphylogeneticspeciesofpossiblyhybrid origins,sistertoacladeincluding>20 Salmo species[12].
InnorthernItaly,northernAdriatic S.marmoratus populationsarestronglyassociated withthe“Marmoratus”(MA)haplogroupofthemtDNAcontrolregion(D-loop)[22,69–72]. However,MAhaplotypeshavealsobeenfoundinseveralbrowntrouttaxaandpopulations ofGreece,Albania,Croatia,centralItaly,andCorsica,e.g.,[23,48,66,71].Likeseveral otherbrowntrouttaxaandpopulations,intheBalkansSouthernAdriatic S.marmoratus populationscanbeassociatedwiththe“Adriatic”(AD)mtDNAhaplogroup[67,72].
MitochondrialmolecularclocksestimatedmuchmorerecentoriginsoftheMAand ADhaplogroups(0.21–0.05myaand0.39–0.13mya,respectively,consideringthe95% highestprobabilitydensityintervalsestimatedusingtwodifferentsubstitutionrates[23]) thanthetimeofdivergencebetween S.marmoratus andthenuclear“S.trutta complex”.It wassuggestedthattheobservedmitochondrial-nuclearphylogeneticdiscordancemightbe theeffectofincompletelineagesortingorasymmetricintrogressivehybridization(mtDNA capture;e.g.,[73]).Themucholdertimeofdivergencebetweentheselineagesrelative tothetimeofhaplogroupdifferentiationstronglysupportsthelattermechanism.PaleointrogressivehybridizationbetweenthemarbletroutandtheApennineMediterranean troutcouldhaveoccurredduringseveralsecondarycontactsasaconsequenceofthe expansionofthePopaleo-basinduringglacialmaxima,asitoccurredinother Salmo species[22,32,53,59,66,74].MosaicdistributionsofmtDNAhaplogroupsamongdifferent taxaarecommonalsoinareaswithoutahistoryofnon-nativetrout’sstocking(e.g.,Albania[75]),andsimilardistributionalanddiversitypatternsmighthaveoccurredinItaly aftertheLastGlacialMaximum(LGM~18,000yearsago).
Phylogeneticpatterns,molecularclocksandthezoogeographyofcongenerssuggestthat S.marmoratus isoneofthe Salmo lineagesthatdivergedinthepaleo-Adriatic drainage,infreshwaterrefugesformedduringtheprecedingLagoMarephase(~5mya). DuringthePleistocene,reducedsalinity,coolerseatemperatures,andextensivepalaeoriverbasinswouldhavefacilitatedthewestwarddispersalofthesefreshwaterlineages acrosstheregionthroughmultiplewavesofcolonization,bottlenecks,andsecondary contacts [32,72,75],allowing S.marmoratus tocolonizetheorographiclefttributariesof thepalaeo-Pobasin [56,76,77].AftertheLGM,increasedsalinitylevelsandsea-levelrise disconnectedthesepopulations,facilitatingallopatricfragmentationanddifferentiationof mtDNAlineages,resultinginthepresentgeographicdistribution[32].
InthenorthernAdriaticbasin, S.marmoratus exhibitawest-to-eastgeographicgradient inMA-s1andMA-s2haplotypedistribution,consistentwiththedescribedstepwisewestwardmigrationandphylogeographicscenario[78].Significantmicrogeographicgenetic differentiationwasalsomeasuredwithinbasins,e.g.,betweenriversandtheirtributaries, suggestingthepresenceoflimitedgeneflowamongdifferentpopulations[79,80].Acontact zonebetween S.marmoratus and S.ghigii wasfoundintheSouth-westernAlps(Section 4).
4.Presenceof S.ghigii intheItalianAlpineandSubalpineRegion
WithintheItalianAlpineregion[81],viablenativepopulationsof S.ghigii (Section 2) haveonlybeenfoundintheSouth-westernAlps(CottianandMaritimeAlps:upper SturadiLanzo,upperDoraRiparia,upperChisone,upperPellice,upperPo,upperStura Demonte,upperGesso,andupperTanarobasins),whereacontactzonewith S.marmoratus wasdescribed,alongageographicdistributiongradientofgeneticvariantsassociated withdifferenttroutphyleticlineages[20–23,34,48,50,77,80,82,83];Section 3.TheMA,AD, and“Mediterranean”(ME)haplotypeprobabilitydensitiesrelativetoelevationshowan altitudinalzonationsuggestinglocalhabitatdifferentiationbetweenthetwoparapatric species,with S.marmoratus beingdominantat0–1000mabovesealevel(asl)and S.ghigii at1000–2000masl[23].Thesefindingsareconsistentwiththedistributionalpatternsof troutswithdifferentphenotype,describedinsomehistoricalaccounts[84–86].
TheSouth-westernAlpsareaknownglacialrefuge,wherenativepopulationsof S. marmoratus and S.ghigii couldhavesurvivedtheLGM[82].IntrogressionratesofalienAtlanticgenesintonativetroutpopulationsareherehighlyvariable(0–70%;[20]).Incontrast, inmostoftheNorth-westernandSouth-easternAlps[81]onlythelowertractsoftherivers wereunaffectedbytheicecapduringtheLGM.Assumingthat S.ghigii and S.marmoratus exhibitedahabitatsegregationpatternanalogoustothatpresentlyobservedintheSouthwesternAlps,theLGMlikelyallowedthesurvivalof S.marmoratus atloweraltitudes, while S.ghigii mighthavebeenpushedintothemarbletrouthabitatandoutcompeted[82]. Afterthelastglaciation,mostAlpinelakesandheadwatersmayhaveonlymarginallybeen colonizedby S.marmoratus andlikelyremained“fishless”(i.e.,troutless).Inhistoricaltimes, thesesystemswereartificiallystockedwithtranslocatedsalmonids,includingnon-native Atlantic S.trutta [82],tosupportsubsistenceandrecreationalfisheries[87–89].Thecapacity of S.marmoratus tooutcompeteothertroutspecieswasobservedbySommani[90],who observedthatinspecificwatercoursesmarbletroutisabletorapidlyreplacebrowntrout (S.truttafario = S.ghigii or S.trutta;thisauthorwasunabletodiscriminatebetweenthese species),whenrestockingpracticesareinterrupted.
IntheLakeofGardabasin,aknownglacialrefugium[91]intheSouth-eastern Alps[81],astudy[92]foundtracesofthemitochondrialvariantADcs-1(themostwidespread ADhaplotype[23],typicallyassociatedwiththe“Adriaticgrouping”of S.truttafario, sensu[50]= S.ghigii).Theprehistoricpresenceof S.ghigii intheLakeGardarefugium isalsoconsistentwiththepresencein S.carpio ofhaplotypesphyleticallyrelatedtohaplogroupstypicallyassociatedwith S.ghigii (AD)and S.marmoratus (MA),suggesting thatoneormorepaleohybridizationeventsoccurredinthisbasinbetweenthesetrout lineages[20,23,50,53].Thisalsosupportsthehypothesisofextensivesecondarycontacts andhybridizationeventsbetweenpeninsularandmarbletroutlineagesbeforethelast glaciationinthisregion[53].TheADcs-1haplotypewasalsofoundintwomuseumspecimenswithlacustrinemorphscollectedinLakeGardaandLakeMaggiorein1877and 1879,respectively[83].LakeMaggioreisanotherknownglacialrefugium[91,93]located intheNorth-westernAlps[81].ThepresenceofADhaplotypesinthesebasinssuggests thatrelictpopulationsof S.ghigii mighthavesurvivedtheLGMinotherglacialrefugia oftheNorth-westernandSouth-easternAlps.Morespeculatively,since S.marmoratus is theonlynativetroutwithlacustrinemorphsinthisregion,thismightalsoindicatethe morecommonpresenceofmarbletroutswithADhaplotypesinthisbasininpasthistorical times,oreventhepresenceofrecentlyextinctandundescribedtrouttaxa[20,94,95].
WiththeonlyexceptionoftheSouth-westernAlps,theabsenceofviablepopulations of S.ghigii innorthernItalyclearlyindicatesthatallrelictpopulationsof S.ghigii thatmight havesurvivedtotheLGMinotherglacialrefugiaweresubsequentlyextirpated.This mighthavereasonablyoccurredduetodemographicorgeneticswamping[96]causedby theman-mademassiveandprolongedintroductionsofnon-native S.trutta inhistorical times.Consistently,introgressionratesofAtlantic S.trutta into S.marmoratus arehigher intheNorth-westernandSouth-easternAlps,andonlytracesofthehaplogroupstypicallyassociatedwithnative S.ghigii werefound[21,82].Thisscenarioissupportedby theprobabilisticapproachadoptedby[92],whichshowedthat,inspiteofthemassive introductionsof S.trutta,genetictracesofextirpated S.ghigii populationscouldstillbe foundinsomeglacialrefugialareassuchastheLakeGardabasin.Suchdramaticeffects couldhavebeenfacilitatedbystrongnumericaldifferencesbetweennative S.ghigii populationsand S.trutta introductions,lowhybridfitness,andweakreproductivebarriers. Bycontrast,thepresenceofpartialreproductivebarriersbetweennon-native S.trutta and S.marmoratus [80,97],thecompetitiveadvantageof S.marmoratus [90],andmarble-trout stockingcouldhavepreventedthelineageorlocalgenomicextinctionofthelatterspecies. Inspiteofthepresenceofhighintrogressionrates[82],neitherdemographicswamping norlocalgenomicextinctionsofnativeApennine S.ghigii haveeverbeendescribedinthe Tuscano-LatiumItalianichthyogeographicregion,wherenon-native S.trutta havebeen andarebeingintroduced.Ontheotherhand,these S.ghigii populationsweremuchless impactedbyhabitatmodificationorcompetitionwithotherspeciesduringtheLGM,and werelikelylargerandlessfragmentedwhentheywerefloodedby S.trutta introductions. SomestudiesfoundtheallozymicvariantsLDH-C1*100andTF*102,typicallyassociatedto S.ghigii populationsintroutpopulationsnativetoFranceandsouth-west Piedmont,insitescollectedeastoftheSouth-westernAlps,hencesuggestingthepresence of S.ghigii outsidethementionedcontactzonewith S.marmoratus [23].However,these allozymeswerealsofoundathighfrequencyinDanubiannativepopulationsofdifferent Salmo species[49,98,99].Inoneofthesestudies,LargiadèrandScholl[100]assumedthe nativestatusofan“Adriaticfario”inalargeportionofthePobasin,basedonmolecular studiesconductedinsouth-westernPiedmont[49]andonphenotypicstudiesthatwere howeverunabletodiscriminatebetweenAtlantic(S.trutta)andAdriatic(S.ghigii)trout phenotypes[101].Theseauthorsfoundthesetwoallozymicvariantsathighfrequency (~20–30%)inEngadin(Danubianbasin),intheMüstair,tributaryoftheAdigeRiver,and inthePoschiavovalley(Pobasin,PoschiavinoTorrent,tributaryoftheAddaRiver);and atlowfrequency(~0–10%)intheTicinoandValaisbasins,includingatributaryofthe DiveriaTorrent(Chrummbach).No“purebred”individualswerefound.Infact,allthese populationshadbeendirectlyorindirectlyeitherentirelyreplacedorheavilystockedwith troutlineagesoftheDanubianbasinviathePoschiavohatcheries,foratleastonecentury beforethestudycollection[100].Thiswouldexplainthegeneticsimilaritiesbetweenthe troutsofthePoschiavinoandTicinovalleys,subsequentlydetectedbyotherstudiesusing microsatelliteandAFLPmarkerstoinvestigatetheadaptivedivergenceandphylogeographicpatternsoftroutpopulationsoftheRhine,Rhone,andPobasins[102,103].Just like[100],alsothesestudiesassumedthepresenceofan“Adriatictrout”(S.cenerinus, sensu[39]= S.ghigii;Section 2)inthePoschiavinoandTicinovalleys,basedontheliterature[39,100,101].However,giventhelackofgeneticreferences(allelesizerange)of Danubiantroutpopulations(possibly Salmolabrax Pallas1814[102,103])itisnotpossibleto knowwhethertheobserved“Adriatic”genetictracesinthePoschiavinoandTicinovalleys wereoriginallypresentinthisregion,orwereleftbyintroducedDanubianstocks[102], asalsosuggestedbythepresenceofDanubianmtDNAhaplotypes(DAhaplogroup)in theTicinobasin[78].Kelleretal.[102,103]alsofoundevidenceofintrogressionofthe PoschiavopopulationintooneRhinepopulation(SE).SEistheclosestRhinepopulationto theDanubiandrainage,suggestingthepresenceofstockingactivitiesandtranslocations betweenSE,Poschiavo,andDanubiansystems.
of Danubian trout populations (possibly Salmo labrax Pallas 1814 [102,103]) it is not possible to know whether the observed “Adriatic” genetic traces in the Poschiavino and Ticino valleys were originally present in this region, or were left by introduced Danubian stocks [102], as also suggested by the presence of Danubian mtDNA haplotypes (DA haplogroup) in the Ticino basin [78]. Keller et al. [102,103] also found evidence of introgression of the Poschiavo population into one Rhine population (SE). SE is the closest Rhine population to the Danubian drainage, suggesting the presence of stocking activities and translocations between SE, Poschiavo, and Danubian systems.
There are several descriptive accounts (cuisine recipes, anecdotes, poetry, and even paintings e.g., [104]) of trouts in the North-western and South-eastern Alps (e.g., Lakes of Como and Garda basins) before the dramatic expansion of the fish-culture industry that promoted the rapid diffusion of the non-native Atlantic S. trutta in the early 19th century (1850–1893, [10]). Several ones [105–112] depict or describe trouts without a marbled coloration pattern and with either red and black dots, phenotypically compatible with several trout taxa, or with speckled dark patterns on a silvery background, compatible with a generalized pelagic (lacustrine) morph of anadromous trout. Adult S. marmoratus living in rivers typically exhibit a marbled coloration pattern [113]; however, anadromous individuals in pelagic conditions can exhibit a silvery and dark-speckled coloration pattern, even leading to taxonomic confusion, e.g., [114].
Thereareseveraldescriptiveaccounts(cuisinerecipes,anecdotes,poetry,andeven paintingse.g.,[104])oftroutsintheNorth-westernandSouth-easternAlps(e.g.,Lakesof ComoandGardabasins)beforethedramaticexpansionofthefish-cultureindustrythat promotedtherapiddiffusionofthenon-nativeAtlantic S.trutta intheearly19thcentury (1850–1893,[10]).Severalones[105–112]depictordescribetroutswithoutamarbled colorationpatternandwitheitherredandblackdots,phenotypicallycompatiblewith severaltrouttaxa,orwithspeckleddarkpatternsonasilverybackground,compatible withageneralizedpelagic(lacustrine)morphofanadromoustrout.Adult S.marmoratus livinginriverstypicallyexhibitamarbledcolorationpattern[113];however,anadromous individualsinpelagicconditionscanexhibitasilveryanddark-speckledcolorationpattern, evenleadingtotaxonomicconfusion,e.g.,[114].
Youngmarbletrouttypicallyexhibitanirregularblackorred-and-blackdottedpattern, withalargeblackpreopercularblotch,similartoadultbrowntrouts[113](Figure 2a); thedarkdottedpatterncanchangetoamarbledpatterninafewmonthsinsubadults (Figure 3a–d);andadultslivinginsmallandfast-flowstreamscanbecomereproductiveat halfthetypicallengthatmaturity,whileretaininga“brown-trout”redorred-and-black dottedpattern[115](Figure 2b,c).
Young marble trout typically exhibit an irregular black or red-and-black dotted pattern, with a large black preopercular blotch, similar to adult brown trouts [113] (Figure 2a); the dark dotted pattern can change to a marbled pattern in a few months in subadults (Figure 3a–d); and adults living in small and fast-flow streams can become reproductive at half the typical length at maturity, while retaining a “brown-trout” red or red-and-black dotted pattern [115] (Figure 2b,c).

Figure2. Examplesofdottedcolorationpatternsin S.marmoratus:juvenilecolorationpattern (a),andadultslivinginsmallandfast-flowingstreams(b,c);(a)juvenilefromRoledo(Piedmont, Verbano-Cusio-Ossola:VCO;46◦ 10 16.7 N8◦ 18 49.7 E),15.5cmtotallength—TL,29.0gwetmass, 22months ofage,black-and-reddottedpattern,MAhaplogroup,qMa 0.995(admixtureproportionof aclusterincludingpurebred S.marmoratus references),90%BCI0.966–1.000;(b)adult(reproductive) specimenfromRioIschielle,tributaryoftheAvisioTorrent(ProvinceofTrento);thespecimenwas collectedfromapopulationwhichresidedfor2generationsinthissmallstream,whichdescended fromhatchery-reared S.marmoratus withmarbledphenotypecollectedfromtheAdigeRiver[115]; 26.9cmTL,courtesyofLeonardoPontalti;(c)adult(reproductive)specimenfromRiodellaBalma, tributaryoftheSangoneRiver(ProvinceofTorino),18.5cmTL,MAhaplogroup,qMa 0.996,90%BCI 0.978–1.000,courtesyofPaoloLoConte.
Biology 2022, 11,576
bano-Cusio-Ossola: VCO; 46°10′16.7″ N 8°18′49.7″ E), 15.5 cm total length—TL, 29.0 g wet mass, 22 months of age, black-and-red dotted pattern, MA haplogroup, qMa 0.995 (admixture proportion of a cluster including purebred S. marmoratus references), 90% BCI 0.966–1.000; (b) adult (reproductive) specimen from Rio Ischielle, tributary of the Avisio Torrent (Province of Trento); the specimen was collected from a population which resided for 2 generations in this small stream, which descended from hatchery-reared S. marmoratus with marbled phenotype collected from the Adige River [115]; 26.9 cm TL, courtesy of Leonardo Pontalti; (c) adult (reproductive) specimen from Rio della Balma, tributary of the Sangone River (Province of Torino), 18.5 cm TL, MA haplogroup, qMa 0.996, 90% BCI 0.978–1.000, courtesy of Paolo Lo Conte.

Figure 3. Examples of dotted coloration patterns in S. marmoratus: conspicuous ontogenetic chromatic variation in pit-tagged individuals which were recaptured at different times; (a,b) subadult specimen sampled in Roledo (Piedmont, Verbano-Cusio-Ossola: VCO; 46°10′16.7″ N 8°18′49.7″ E), age and genetic data unavailable; (a) sampled on 28 April 2021, 20.0 cm TL, 86 g, dotted pattern; (b) recaptured in the same site on 28 October 2021, 23.4 cm TL, 122 g, marbled pattern; (c,d) Subadult specimen sampled in Prata di Vogogna (Piedmont, VCO; 46°1′40.8″ N 8°17′2.2″ E), age and genetic data unavailable; (c) sampled on 26 April 2021, 20.6 cm TL, weight not available, dotted pattern; (d) recaptured in the same site on 19 October 2021, 26.4 cm TL, 166.0 g, marbled pattern.
Figure3. Examplesofdottedcolorationpatternsin S.marmoratus:conspicuousontogeneticchromaticvariationinpit-taggedindividualswhichwererecapturedatdifferenttimes;(a,b)subadult specimensampledinRoledo(Piedmont,Verbano-Cusio-Ossola:VCO;46◦ 10 16.7 N8◦ 18 49.7 E), ageandgeneticdataunavailable;(a)sampledon28April2021,20.0cmTL,86g,dottedpattern; (b)recapturedinthesamesiteon28October2021,23.4cmTL,122g,marbledpattern;(c,d)Subadult specimensampledinPratadiVogogna(Piedmont,VCO;46◦ 1 40.8”N8◦ 17 2.2”E),ageandgenetic dataunavailable;(c)sampledon26April2021,20.6cmTL,weightnotavailable,dottedpattern; (d)recapturedinthesamesiteon19October2021,26.4cmTL,166.0g,marbledpattern.
Ontheotherhand,thereisampleevidenceofsalmonidintroductionsinoldhistorical timesfromoutsideItaly.Domesticationpracticesandtranslocationsoffreshwaterfishes, evenacrossmountainranges,gobacktotheMiddleAgesandpossiblytotheNeolithic, seamlesslycontinuingthroughtothe18thand19thcentury,beforetheonsetofthefishcultureindustry[10,116–119].Non-nativetroutswith“brown-trout”dottedcoloration patternscouldhavebeenintroducedinnorthernItalyfromadjacentareassuchasthe orographicrighttributariesofthePoRiver,orevenbeyondtheAlpineDivide,e.g.,from theDanubebasin,suchasthecommoncarp Cyprinuscarpio L.intheRomanPeriod[10,120].
Considering(i)theabsenceofviablepopulationsof S.ghigii innorthernItaly,except intheSouth-westernAlps(Figure 4a),andthepresenceofpotentialgenetictracesof extirpatedpopulationsinglacialrefugia(Figure 4b–e);(ii)thepastwidespreadpresenceof S.ghigii inthisregionbeforeLGM,supportedbytheestimatedgeneflowoccurredfortens ofthousandsofyearsbetweenthemarbleandpeninsularlineages[53];(iii)thehypothetical extirpationofmostnativepopulationsof S.ghigii inthisregionduringLGM[82];(iv)the possibilityofrecentextirpationsof S.ghigii populationsduetogeneticordemographic swampingcausedbymassiveintroductionsofnon-native S.trutta duringthelasttwo centuries[92];(v)theanecdotalaccountspotentiallyreportingthepresenceof S.ghigii inthisregioninhistoricaltimes,4–5centuriesbeforethe19th-centuryflourishingofthe
fish-cultureindustry[104–112];fourpotentiallyfalsifiablescenarioscanbehypothesized (Figure 4):
1. Nativepopulationsof S.ghigii havebeenextirpatedinmostoftheregionduringthe LGM,exceptinglacialrefugia. S.ghigii wasneversubsequentlyintroducedfrom areasoutsideitsoriginaldistribution.Thentheserelictnativepopulations,e.g.,the glacialrefugiaofLakeMaggioreandLakeGarda,havebeenextirpatedbygeneticor demographicswamping,duetothemassiveintroductionsof S.trutta,exceptthoseof theSouth-westernAlps(Figure 4b).
2. Nativepopulationsof S.ghigii havebeenextirpatedinmostoftheregionduring theLGM,exceptinglacialrefugia. S.ghigii wassubsequentlyintroducedfrom areasoutsideitsoriginaldistribution.Thenboththerelictnativeandnon-native populationshavebeenextirpatedbygeneticordemographicswamping,duetothe massiveintroductionsof S.trutta,exceptthoseoftheSouth-westernAlps(Figure 4c).
3. Nativepopulationsof S.ghigii survivedtheLGMinseveralareasoftheregion, includingglacialrefugia. S.ghigii wasneversubsequentlyintroducedfromareas outsideitsoriginaldistribution.Thenthesenativepopulationshavebeenextirpated bygeneticordemographicswampingduetothemassiveintroductionsof S.trutta, exceptthoseoftheSouth-westernAlps(Figure 4d).
4. Nativepopulationsof S.ghigii survivedtheLGMinseveralareasoftheregion, includingglacialrefugia. S.ghigii wassubsequentlyintroducedfromareasoutsideits originaldistribution.Thenboththerelictnativeandnon-nativepopulationshavebeen extirpatedbygeneticordemographicswamping,duetothemassiveintroductionsof S.trutta,exceptthoseoftheSouth-westernAlps(Figure 4e).
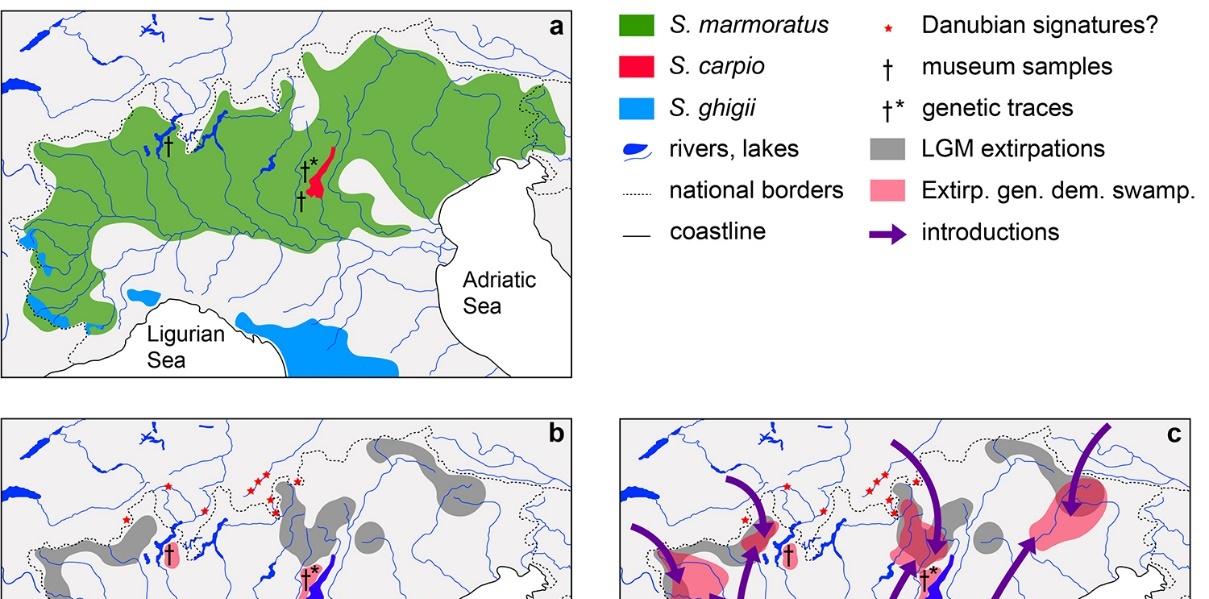
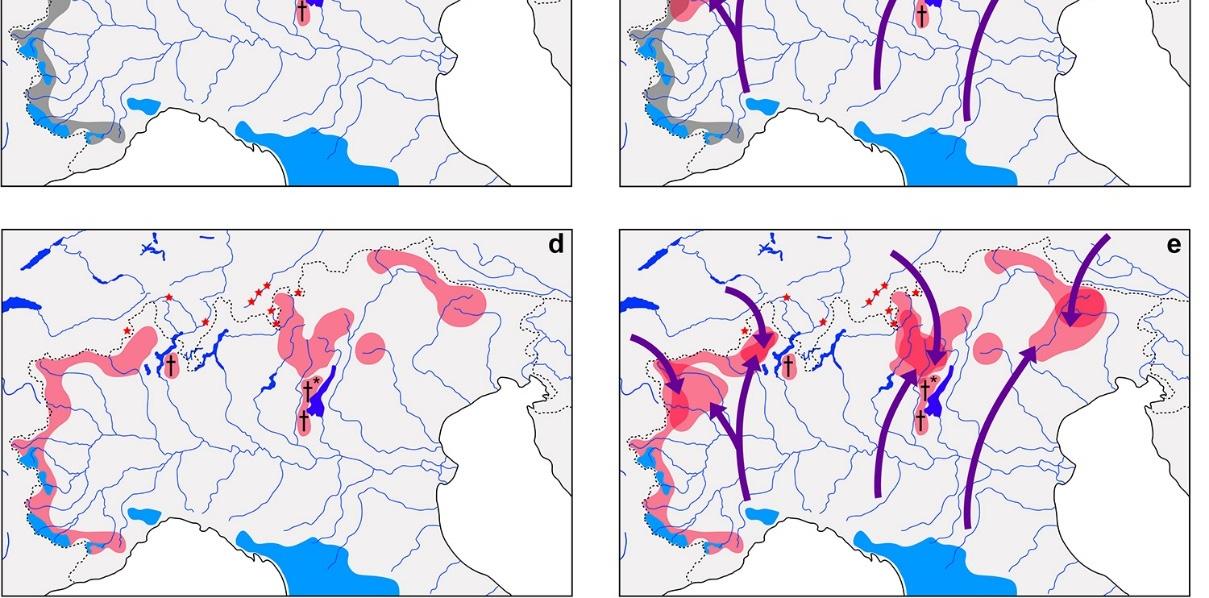
Figure4. PresentdistributionoftroutsinnorthernItalyandhypotheticalreconstructionsofthe
Figure 4. Present distribution of trouts in northern Italy and hypothetical reconstructions of the demographic history of S. ghigii in this geographic area; (a) present distribution of Alpine and subalpine trouts in northern Italy, modified from [23,48,90,121]; (b) graphical representation of the hypothetical distribution of S. ghigii that were extirpated during the LGM except in glacial refugia, lack of subsequent introductions of S. ghigii in historical times, and subsequent extirpations of relict native populations in glacial refugia (e.g., Lake Maggiore, Lake Garda, except the South-western Alps), due to genetic or demographic swamping caused by the massive introductions of S. trutta during the last ~150 years; (c) as in (b), but with man-made introductions of S. ghigii from areas outside its
demographichistoryof S.ghigii inthisgeographicarea;(a)presentdistributionofAlpineand subalpinetroutsinnorthernItaly,modifiedfrom[23,48,90,121];(b)graphicalrepresentationof thehypotheticaldistributionof S.ghigii thatwereextirpatedduringtheLGMexceptinglacial refugia,lackofsubsequentintroductionsof S.ghigii inhistoricaltimes,andsubsequentextirpations ofrelictnativepopulationsinglacialrefugia(e.g.,LakeMaggiore,LakeGarda,excepttheSouthwesternAlps),duetogeneticordemographicswampingcausedbythemassiveintroductionsof S.trutta duringthelast~150years;(c)asin(b),butwithman-madeintroductionsof S.ghigii from areasoutsideitsoriginaldistribution;(d)hypotheticaldistributionof S.ghigii populationsthat survivedtheLGM,lackofsubsequentintroductionsof S.ghigii inhistoricaltimes,andsubsequent extirpationsofrelictnativepopulationsinglacialrefugia,excepttheSouth-westernAlps,due togeneticordemographicswampingcausedbythemassiveintroductionsof S.trutta;(e)asin (d),butwithman-madeintroductionsof S.ghigii fromareasoutsideitsoriginaldistribution.In legend,presentdistributionofthethreetroutspecies: S.marmoratus (greenarea), S.carpio (redarea), and S.ghigii (bluearea);Danubiansignatures?=recordsofgeneticvariantspossiblyintroduced fromtheDanubebasin[100,102,103];museumsamples=recordsofADcs-1intwo19th-century museumsamples[83];genetictraces=potentialtracesofextirpated S.ghigii populations[92];LGM extirpations=hypotheticaldistribution(greyarea)of S.ghigii populationsthatwereextirpatedduring LGM;Extirp.gen.demswamp.=hypotheticaldistribution(pinkarea)of S.ghigii populations thatwereextirpatedbydemographicorgeneticswampingcausedbyintroductionsof S.trutta in historicaltimes;introductions=directions(purplearrows)of S.ghigii man-madeintroductionsfrom areasoutsideitsoriginaldistribution.
Ineveryscenario, S.ghigii hasbeenextirpatedinthisregionexceptintheSouthwesternAlps,andnoothernativeandviablepopulationsareleftatpresent.Thepopulationsof S.ghigii oftheSouth-westernAlpsaretheonlyautochthonousones,therefore beingtheonlyonesthatmightbemanaged.
Inboththefirstandsecondscenarios(Figure 4b,c) S.ghigii becamenon-nativeinthe AlpineandsubalpineItalianregionaftertheLGM,exceptinglacialrefugia,asindicatedby theavailablescientificevidence.Duetotheintenseclimaticandecologicalchangesthat followedtheLGM,itwouldmakelittleconservationalandecologicalsensetoreintroduce speciesorpopulationsthatlivedinnorthernItalyduringtheLatePleistoceneandwere naturallyextirpatedduringtheLGM,e.g.,suchastheleopard Pantherapardus L.[122,123]. ExcepttheSouth-westernAlps,populationsof S.ghigii mayhavesurvivedinotherglacial refugia,beforebeingextirpatedbydemographicorgeneticswampingcausedby S.trutta introductionsinhistoricaltimes.Evenifthesereconstructionsaresupportedbyfurther investigations,anyreintroductionof S.ghigii insuchareas(e.g.,LakeMaggioreandLake Gardabasins)shouldtakeintocarefulconsiderationthepresentenvironmentalconditions, potentialinteractionswithinthecommunity,andhabitatavailability,sincethesemayhave becomeinadequatetohostthespeciessinceitsextirpation.Thiscaseisexemplifiedby theEurasianbeaver(Castorfiber L.),whosenorth-Italianpopulationssurvivedinglacial refugiaduringtheLGMandsubsequentlyre-expandedintheregion,onlytobecompletely extirpatedinthe16th–17thcenturyduetoanthropogenicenvironmentalchanges[124]. InspiteofthesporadicsightingsoftheEurasianbeaverinnorthernItalyin2018and 2020[125],thelackofavailablehabitatinthisregionwouldlikelymakeareintroduction programmeaningless.
Inthethirdandfourthscenarios(Figure 4d,e),nativepopulationsof S.ghigii would havesurvivedtheLGMinseveralareaswithinthisregion,includingglacialrefugia, andwereeventuallyextirpatedbydemographicorgeneticswampingcausedbythe introductionsof S.trutta.Thesescenarioslackscientificsupport,sincethepresenceof potentialtracesof S.ghigii outsideglacialrefugiawasneverdemonstrated.However, evenifthepastpresenceof S.ghigii inotherareasisdemonstratedinthefuture,the sameconsiderationsmadeforscenarios1and2apply,incaseofextirpationsoccurredin historicaltimes.Carefulassessmentsofenvironmentalimpactmustbeconducted,before consideringanyreintroductionsof S.ghigii insuchareas,irrespectiveofitsstatuspreceding
itsextirpation.Thereforeintheabsenceofreliabledata,reintroductionsof S.ghigii should beavoided,invokingthePrecautionaryPrinciple[126].
Notwithstanding,massiveamountsofnon-nativeindividualsof S.ghigii sourced fromoutsidethisregionhavebeenandarebeingregularlyandmassivelyintroducedinto thisregionduringthelastdecade.Suchstockingactivitieswillobviouslyhamperany investigationattemptingtoassessthestatusofanyrelict S.ghigii populationsthatmaybe foundinthisregioninthefuture.Infact,anynewbiologicalvariantfoundintheregion thatisnotpresentinanyofthenon-nativesourcepopulationsmaystillbenon-native,i.e., beingstillundescribedinthesourcepopulation.
Theriskofintrogressivehybridizationbetweennative S.marmoratus andnon-native stocksof S.ghigii isnotonlysuggestedbythewidespreadinterfertilitybetween Salmo species[12,72].Thesamepresenceofgene-flowbetweenthesetroutlineagesduringpast secondarycontactsbeforeLGM[53]clearlydemonstratesthepotentialforsuchevents. Thisisparticularlythecasewhennativesmallandfragmentedpopulationsarefloodedby largeamountsofstockednon-nativefish[82],inthesameecologicalconditionsthatcaused theongoingintrogressionbetween S.marmoratus andAtlantic S.trutta inthisregion[127].
Theongoingstockingofindividualsof S.ghigii obtainedfromnon-nativepopulations isalsoassociatedwithotherrisks.Whilehatcherymanagersintroducing S.ghigii in thisregiontypicallydonotpublishanygeneticscreeningofthestocks,anindependent investigationinadifferentgeographicareashowedthatahatcherystockof“Mediterranean trouts”actuallycontainedamixofbothAtlantichaplotypes(44%)andMediterranean haplotypes(66%),beingcharacterizedbyaqAT value(admixtureproportionofthecluster characterizingtwohatcherystocksofnon-nativeAtlantic S.trutta)of0.42–1.00[19].Given theexceptionalrarityofsuchindependentinvestigations,suchcasesaremorelikelythe ruleratherthantheexception.Introductionsofsuchhybridstocksposeanevengreater riskthanintroductionsofpurebrednon-native S.ghigii,sincehybridscaneffectivelyact asageneticbridge,facilitatinghybridizationandintrogressionbetweenreproductively isolatedspecies[128].
Thedisputerevolvingaround S.ghigii andthe“Mediterraneantrout”innorthernItaly isalsorelatedtoanotherhotandcurrenttopic,i.e.,themanagementofstreamheadwaters. Stockingofnative S.marmoratus generallyoccursinwhatisconsideredtobeitsputative vocationalhabitat,i.e.,inmiddleandlowerriverreaches(<1500masl[90]).Ontheother hand,currentregulationshaveoftenprohibitedtheintroductionofnon-nativespecies, suchastheAtlanticbrowntrout,thathastraditionallybeenreleasedinthesewatercourses for>150years(Section 1).Thiscausedaheateddebate,fueledbyanglersandsportfishing associations,whoadvocatetheuseofnon-nativestocksof S.ghigii or S.trutta toexploit streamheadwaters.Priortohumansettlementorstocking,mostheadwaterstreamsinthis regionwerelikelytroutless(Section 4).However,whilethishypothesisstilllacksexperimentalsupport,theideaofstockingnon-native S.ghigii intheseenvironmentsisassociated withsubstantialenvironmentalrisk,beingatoddswithconservationprinciples(Sections 1 and 4).Headwaterstreamscouldbestockedwithnon-nativesterilizedfishes,e.g.,bya processoftriploidization,thuspreventinghybridizationwithnativespeciesdownstream. However,massivefishintroductionscansignificantlyimpactthestructureandfunctioning offreshwaterecosystems,duetointeractionswiththenativecommunitiesandrecipient environments,includingincreasedcompetition,predation,biogenicmodificationofthe environment,andpotentialspreadingofdiseases[129–134].Therefore,thesustainability ofthismanagementstrategymustbeconsideredwithextremecaution,carefullyassessed, andregularlymonitoredafterimplementation.
6.ConclusionsandFutureDirections
Atpresent,robustscientificevidenceidentifies S.marmoratus plussmallandfragmentedpopulationsof S.ghigii intheSouth-westernAlpsastheonlynativeanddistinct
Salmo lineagesandpopulationsinnorthernItaly.Introgressivehybridizationfromnonnative Salmotrutta into S.marmoratus inthisregionhasbeenrepeatedlydemonstrated. Giventheevidenceofpastintrogressionfromthepeninsularlineageintothemarblelineage, geneticintrogressionfromnon-nativestocksof S.ghigii intonative S.marmoratus mayhave alreadyoccurred.Inparticular,thepresenceinnon-nativeintroducedstocksof S.ghigii of life-historytractsthatwouldpreventorminimizehybridizationwith S.marmoratus cannot andshouldnotbeassumed.
Possiblegenetictracesof S.ghigii outsidethecontactzonearefragmentary,and stronglysuggestpastextirpations.Itispresentlyunknownwhetherviablepopulations ofthisspecieswerepresentinhistoricaltimes,ormostofthenativepopulationswere extirpatedduringtheLGM.
Regardlessoftheacademicvalueoftaxonomicdebates,thehighinterfertilityamong manysalmonidtaxamakesarationalapproachtotheecosystemmanagementofsalmonid populationsextremelychallenging.Anyintroductionoftroutindividualsoriginating fromnon-nativepopulationsdefinedbyphylogeographicandgeneticcriteria,evenif conspecificwiththerecipientpopulation,posestheriskofgeneratinghybridswarms betweennon-nativestocksandnativetroutlineages.Forthisreason,translocatedand restockedsalmonidpopulationsshouldalwaysbemanagedasEvolutionarySignificant Units(ESUs;[32,135,136]).Consideringthegeneticstructureofpopulationsatthemicrogeographicscale(i.e.,hydrogeographicbasinandsub-basin)makesconservationactions taxonomy-independent,gainingthesorelyneededstabilityforconservationpurposes. Inthespecificcaseofreintroductions,usingtheclosestavailableESUasasourcetorebuildanextirpatedpopulationstillposestheriskofintroducingindividualswithdifferent life-historytraitsthanthoseoftheoriginalpopulation.Forthisreason,thepotentialimpactsofreintroductionsshouldalwaysbecarefullyevaluated,incaseprevented,andthen monitored,e.g.,[137](Sections 4 and 5).
Futureresearchcould(i)furtherourknowledgeofthegeneticstructureandmicrogeographicpatternsof S.marmoratus populationswithintheItalianAlpineandsubalpine region,thusidentifyingESUsforscience-basedconservationandmanagement;(ii)investigatethepastpresenceof S.ghigii populationsinthisregionusingancientDNA,e.g., inarchaeologicalsites[138]orinancientlakesediments[139];(iii)investigatethepast presenceofsalmonidsinhypotheticallytroutlessheadwaterstreamspriortostocking, e.g.,usingmuseumrecords;(iv)investigatethepresenceofvertebrateandinvertebrate speciesofevolutionaryandconservationinterestevolvedinhypotheticallytroutlessheadwaterstreams,andtheirpotentialinteractionswithnon-nativefishcandidatesforstocking; (v)monitor spatiotemporaldynamicsofgeneticdiversityoftroutlineages,bothnative andnon-native,withspecialattentiontothepotentialonsetanddevelopmentofnew hybridswarmsoriginatingfromtherecentintroductionsfrom S.ghigii non-nativestocks; (vi)investigatethegeneticstructure,ecologyandconservationstatusof S.ghigii native populations(ESUs)intheSouth-westernAlps,designingandimplementingdedicatedconservationprograms,ifneeded;(vii)evaluateandimplementsupportingbreedingprograms forthreatenedandheavilyfishedESUsof S.marmoratus,usingstate-of-the-artmethodsto geneticallyselectbreedersandminimizedomesticationeffects.
Researcheffortswillhoweverbeinsufficienttomeetthecommongoalofsustainable ecosystemmanagement[140],ifallthestakeholdersarenotinvolvedinadurable,empathetic,andcollectiveeffort.Fromanglerstosportfishingassociations,hydropowersectors, researchers,conservationists,andgovernmentalinstitutions,everyoneiscalledtoprotect, conserve,andpromotethenativenorth-Italiantrouts,hencepreservingthenaturalheritage ofourinlandwaters.
AuthorContributions: Conceptualization,G.P.andP.V.;writing—originaldraftpreparation,G.P.; writing—reviewandediting,G.P.,T.R.,M.I.andP.V.;supervision,P.V.;projectadministration,P.V.; fundingacquisition,P.V.Allauthorshavereadandagreedtothepublishedversionofthemanuscript.
Funding: ThisstudywasfundedbyINTERREGITA-CHSHARESALMOandLIFE15NAT/IT000823 IdroLIFEProjects.
InstitutionalReviewBoardStatement: Notapplicable.
InformedConsentStatement: Notapplicable.
DataAvailabilityStatement: Notapplicable.
Acknowledgments: TheauthorswouldliketothankLeonardoPontalti(Ufficiofaunisticodella ProvinciadiTrento)andPaoloLoConte(Città MetropolitanadiTorino)forthephotosof S.marmoratus fromRioIschielleandSangoneRiver,respectively.WealsothankAndreaGandolfi(Fondazione EdmundMach),AndreaSplendiani(Univ.PolitecnicadelleMarche),LeonardoPontalti,Antonino Duchi(LegambienteRagusa),PaoloLoConte,CesarePuzzi(G.R.A.I.A.srl),GianMauroBertoiaand UmbertoGrossi(F.I.P.S.A.S.VCO),PierpaoloGibertoni(M.T.R.G.),ValterBianchini(U.P.S.Sondrio), LuigiGuglielmetti(A.P.S.Como),GiancarloPerosino(C.R.E.S.T.Piemonte),andthenumerous passionateanglersofnorthernItalymetinthelasttwoyearsforthestimulatingconversations revolvingaroundthistopic.WewouldalsoliketothankAndreaSplendianiforhisvaluablecomments onanearlyversionofthemanuscript.
ConflictsofInterest: Theauthorsdeclarenoconflictofinterest.Thefundershadnoroleinthedesign ofthestudy;inthecollection,analyses,orinterpretationofdata;inthewritingofthemanuscript,or inthedecisiontopublishtheresults.
References
1. Brookes,R. TheArtofAngling,5thed.;LowndesT:London,UK,1781;p.304.
2. Hoffmann,R.C.Troutandfly,workandplay,inmedievalEurope.In Backcasts:AGlobalHistoryofFlyFishingandConservation; Snyder,S.,Borgelt,B.,Tobey,E.,Eds.;UniversityofChicagoPress:Chicago,IL,USA,2016;pp.27–45.
3. Brown,J.C.;Lokensgard,K.H.;Snyder,S.;Draper,M.Theculturalcurrentsandsocialvaluesoftrout.In TroutandCharofthe World;Kershner,J.L.,Williams,J.E.,Gresswell,R.E.,Lobón-Cerviá,J.,Eds.;AmericanFisheriesSociety:Bethesda,MD,USA,2019; pp.65–93.
4. Yu,P.L. Rivers,FishandthePeople:Tradition,Science,andHistoricalEcologyofFisheriesintheAmericanWest;UniversityofUtahPress: SaltLakeCity,UT,USA,2015.
5. Kinsey,D.‘Seedingthewaterastheearth’:Theepicenterandperipheriesofawesternaquaculturalrevolution. Env.Hist. 2006, 11,527–566.[CrossRef]
6. Draper,M.Holytrout:NewZealandandSouthAfrica.In Backcasts:AGlobalHistoryofFlyFishingandConservation;Snyder,S., Borgelt,B.,Tobey,E.,Eds.;UniversityofChicagoPress:Chicago,IL,USA,2016;pp.178–194.
7. Vinciguerra,D.IcoregonielalorointroduzioneinItalia. L’ecoDeiCampiEDeiBoschi 1895, 2,2.
8. Borroni,I.;Grimaldi,E.Fattorietendenzedimodificazionedell’ittiofaunaitalianad’acquadolce. Ital.J.Zool. 1978, 45,63–73. [CrossRef]
9. Bianco,P.G.L’ittiofaunacontinentaledell’Appenninoumbro-marchigiano,barrierasemipermeabilealloscambiodicomponenti primarietraglioppostiversantidell’Italiacentrale. Biogeographia 1994, 17,427–485.[CrossRef]
10. Bianco,P.G.FreshwaterfishtransfersinItaly:History,localmodificationoffishcomposition,andapredictiononthefutureof nativepopulations.In StockingandIntroductionsofFishes;Cowx,I.G.,Ed.;FishingNewBook;BlackwellScience:Oxford,UK, 1998;pp.167–185.
11. Halverson,A. AnEntirelySyntheticFish:HowRainbowTroutBeguiledAmericaandOverrantheWorld;YaleUniversityPress:New Haven,CT,USA,2010.
12. Segherloo,I.H.;Freyhof,J.;Berrebi,P.;Ferchaud,A.-L.;Geiger,M.;Laroche,J.;Levin,B.A.;Normandeau,E.;Bernatchez,L. Agenomicperspectiveonanoldquestion: Salmo troutsor Salmotrutta (Teleostei:Salmonidae)? Mol.Phylogenet.Evol. 2021, 162,107204.[CrossRef][PubMed]
13. OntheConservationofNaturalHabitatsandofWildFaunaandFlora,CouncilDirective92/43/EEC21May,OJL206,22July 1992.pp.7–50.Availableonline: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A31992L0043 (accessedon4 February2022).
14. DecretodelPresidentedellaRepubblica8Settembre1997,n.357,MinisterodellaTransizioneEcologica.Availableonline: https://www.mite.gov.it/ (accessedon14February2022).
15. Ministerodell’AmbienteedellaTuteladelTerritorioedelMare.CriteriperlaReintroduzioneeilRipopolamentodelleSpecie Autoctonedicuiall’allegatoDdelDecretodelPresidentedellaRepubblica8Settembre1997,n.357,eperL’immissionediSpeciee diPopolazioninonAutoctone(20A02112). Gazz.Uff.Repubbl.Ital. 2020, 98,2–6.Availableonline: https://www.gazzettaufficiale. it/eli/id/2020/04/14/20A02112/sg (accessedon9February2022).
16. Lowe,S.;Browne,M.;Boudjelas,S.;DePoorter,M. 100oftheWorld’sWorstInvasiveAlienSpecies:ASelectionfromtheGlobal InvasiveSpeciesDatabase;InvasiveSpeciesSpecialistGroup(ISSG,SpeciesSurvivalCommissionoftheInternationalUnionforthe ConservationofNature):Auckland,NewZealand,2000.
17. PresidentedellaRepubblica.BilanciodiPrevisionedelloStatoperl’annoFinanziario2022eBilancioPluriennaleperilTriennio 2022–2024del31dicembre2021(21G00256). Gazz.Uff.Repubbl.Ital. 2021, 310,1–404.Availableonline: https://www. gazzettaufficiale.it/eli/id/2021/12/31/21G00256/sg (accessedon13February2022).
18. SenatodellaRepubblica(2022)XVIIILegislatura.DisegnodiLeggen.2536.ConversioneinLegge,conModificazioni, delDecreto-Legge30Dicembre2021,n.228,RecanteDisposizioniUrgentiinMateriadiTerminiLegislativi.Available online: https://www.senato.it/japp/bgt/showdoc/18/DDLMESS/0/1334653/index.html?part=ddlmess_ddlmess1&fbclid= IwAR1MYWSt6xKvYr0DUKngfT6Jis0z4oG-b3CE9xE0gTkZOQ86zKAvFY27oQA (accessedon10March2022).
19. Splendiani,A.;Giovannotti,M.;Righi,T.;Fioravanti,T.;Cerioni,P.N.;Lorenzoni,M.;Carosi,A.;LaPorta,G.;CaputoBarucchi,V. Introgressiondespiteprotection:ThecaseofnativebrowntroutinNatura2000networkinItaly. Conserv.Genet. 2019, 65,460–473. [CrossRef]
20. Giuffra,E.;Guyomard,R.;Forneris,G.Phylogeneticrelationshipsandintrogressionpatternsbetweenincipientparapatricspecies ofItalianbrowntrout(Salmotrutta L.complex). Mol.Ecol. 1996, 5,207–220.[CrossRef]
21. Meraner,A.;Gratton,P.;Baraldi,F.;Gandolfi,A.Nothingbutatraceleft?AutochthonyandconservationstatusofNorthern Adriatic Salmotrutta inferredfromPCRmultiplexing,mtDNAcontrolregionsequencingandmicrosatelliteanalysis. Hydrobiologia 2013, 702,201–213.[CrossRef]
22. Splendiani,A.;Palmas,F.;Sabatini,A.;CaputoBarucchi,V.Thenameofthetrout:Considerationsonthetaxonomicstatusofthe Salmotrutta L.,1758complex(Osteichthyes:Salmonidae)inItaly. Eur.Zool.J. 2019, 86,432–442.[CrossRef]
23. Splendiani,A.;Berrebi,P.;Tougard,C.;Righi,T.;Reynaud,N.;Fioravanti,T.;LoConte,P.;Delmastro,G.B.;Baltieri,M.;Ciuffardi, L.;etal.Theroleofthesouth-westernAlpsasaunidirectionalcorridorforMediterraneanbrowntrout(Salmotrutta complex) lineages. Biol.J.Linn.Soc. 2020, 131,909–926.[CrossRef]
24. Gibertoni,P.P.;Pensierini,M.TrotenativedegliAppennini. H2OMag. 2008, 1,56–62.
25. Fricke,R.;Eschmeyer,W.N.;vanderLaan,R.Eschmeyer’sCatalogofFishes:Genera,Species,References.Availableonline: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessedon16January2022).
26. Rafinesque-Schmaltz,C.S. Indiced’IttiologiaSiciliana:Ossia,CatalogoMetodicodeiNomiLatini,Italiani,eSicilianideiPesci,chesi RinvengonoinSiciliaDispostiSecondounMetodoNaturaleeSeguitodaunAppendicecheContienelaDescrizionedeAlcuniNuoviPesci Siciliani;delNoboloG:Messina,Italy,1810.
27. Cuvier,G. LeRègneAnimal,Distribué D’aprèssonOrganisation,pourServirdeBase à l’HistoireNaturelledesAnimauxetd’Introduction à l’AnatomieComparée,2nded.;DétervilleandChrochard:Paris,France,1829;Volume2.
28. Chiereghin,S. Descrizionede’Pesci,de’Crostaceiede’TestaceicheAbitanoleLaguneedilGolfoVeneto;Canovaed:Treviso,Italy,2001; Volume2.
29. Nardo,G.D. SinonimiaModernadelleSpecieRegistratenell’OperaIntitolata:"Descrizionede’Crostacei,de’Testaceiede’Pesciche AbitannoleLaguneeGolfoVenetoRappresentatiinFigure à Chiaro-ScuroedaColori”;Antonelli:Venezia,Italy,1847.
30. Duméril,A.H.A.Notesurunetruited’Algérie(Salarmacrostigma,A.Dum.). C.R.Hebd.Acad.Sci. 1858, 47,160–162.
31. Pomini,F.P.Ricerchesui Salmo dell’Italiapeninsulare.I.LatrotadelSagittario(Abruzzi): Salmoghigii (n.sp.). Atti.Soc.Ital.Sci. Nat.Milano 1941, 80,33–48.
32. Meraner,A.;Gandolfi,A.Geneticsofthegenus Salmo inItaly:Evolutionaryhistory,populationstructure,molecularecologyand conservation.In BrownTrout:Biology,EcologyandManagement;Lobón-Cerviá,J.,Sanz,N.,Eds.;Wiley&SonsLtd.:Hoboken,NJ, USA,2018;pp.65–102.
33. Zerunian,S. IconografiadeiPescidelleAcqueInterned’Italia;Ministerodell’AmbienteedellaTuteladelTerritorio(Direzione ConservazioneNatura).UnionezoologicaItaliana.IstitutoNazionaleFaunaSelvatica.TipolitografiaF.G.SaviglianosulPanaro: Modena,Italy,2002.
34. Bovero,S.;Candiotto,A.;Ceppa,L.;Giuntoli,F.;Pascale,M.;Perosino,G.C.Statodell’ittiofaunaneifiumietorrentidelPiemonte. Riv.PiemontStor.Nat. 2021, 42,135–160.
35. Nardo,G.D. CenniStoricoCriticisuiLavoriPubblicatiSpecialmentenelNostroSecolocheIllustranolaStoriaNaturaledegliAnimali VertebratidellaVenetaTerrafermaedAppendiceRelativaaiTentativiFattinelleProvincieVenetesullaPiscicolturaesullaPropagazione ArtifizialedelPescediAcquaDolce;Grimaldo:Venice,Italy,1875.
36. Heckel,J.J.;Kner,R. DieSüsswasserfischederÖsterreichischenMonarchie,mitRücksichtaufdieAngränzendenLänder;vonEngelmann, W.:Leipzig,Germany,1857.
37. Cuvier,G.;Valenciennes,A. HistoireNaturelledesPoissons.Tomevingtetunième.SuiteduLivrevingtetunièmeetdesClupéoïdes.Livre vingt-deuxième.DelaFamilledesSalmonoïdes;Bertrand,P.,atBerger-Levrault:Strasbourg,France,1848.
38. Kottelat,M.Europeanfreshwaterfishes. Biologia 1997, 52 (Suppl.S5),1–271.[CrossRef]
39. Kottelat,M.;Freyhof,J. HandbookofEuropeanFreshwaterFishes;Kottelat,M.,Freyhof,J.,Eds.;Cornol,SwitzerlandandFreyhof,J.: Berlin,Germany,2007.
40. Snoj,A.;Bravniˇcar,J.;Mari´c,S.;SušnikBajec,S.;Benaissa,H.;Schöffmann,J.NuclearDNArevealsmultiplewavesofcolonisation, reticulateevolutionandalargeimpactofstockingontroutinnorth-westAfrica. Hydrobiologia 2021, 848,3389–3405.[CrossRef]
41. Schöffmann,J.AutochthoneForellen(Salmotrutta L.)inNordafrika. ÖsterreichsFischerei 1993, 46,164–169.
42. Bobbio,L.;Cannas,R.;Cau,A.;Deiana,A.M.;Duchi,A.;Gandolfi,G.;Tagliavini,J.MitochondrialvariabilityinItaliantrouts, withparticularreferenceto“macrostigma”populations.InProceedingsofthe6thNationalConferenceofAIIAD(Associazione ItalianaIttiologiAcqueDolci):CarteItticheDieciAnniDopo,VareseLigure,Italy,6–8June1996;pp.42–49.
43. Duchi,A.Flankspotnumberanditssignificanceforsystematics,taxonomyandconservationofthenear-threatenedMediterraneantrout Salmocettii:Evidencefromageneticallypurepopulation. J.FishBiol. 2017, 92,254–260.[CrossRef]
44. Duchi,A.Extantbecauseimportantorimportantbecauseextant?Onthescientificimportanceandconservationofagenetically pureSicilianpopulationofthethreatened Salmocettii Rafinesque,1810. Cybium 2020, 44,41–44.[CrossRef]
45. Tougard,C.;Justy,F.;Guinand,B.;Douzery,E.J.P.;Berrebi,P. Salmomacrostigma (Teleostei,Salmonidae):Nothingmorethana browntrout(S.trutta)lineage? J.FishBiol. 2018, 93,302–310.[CrossRef][PubMed]
46. Fruciano,C.;Pappalardo,A.M.;Tigano,C.;Ferrito,V.PhylogeographicalrelationshipsofSicilianbrowntroutandtheeffectsof geneticintrogressiononmorphospaceoccupation. Biol.J.Linn.Soc. 2014, 112,387–398.[CrossRef]
47. Duchi,A.;(LegambienteRagusa,Ragusa,Italy).Personalcommunication,2021.
48. Lobón-Cerviá,J.;Esteve,M.;Berrebi,P.;Duchi,A.;Lorenzoni,M.;Young,K.A.TroutandcharofcentralandsouthernEuropeand northernAfrica.In TroutandCharoftheWorld;Kershner,J.L.,Williams,J.E.,Gresswell,R.E.,Lobón-Cerviá,J.,Eds.;American FisheriesSociety:Bethesda,MD,USA,2019;pp.379–410.
49. Giuffra,E.IdentificazioneGeneticaeFilogeniadellePopolazionidiTrotaComune, Salmotrutta L.,delBacinodelPo.Ph.D.Thesis, UniversityofTurin,Turin,Italy,1993.
50. Giuffra,E.;Bernatchez,L.;Guyomard,R.Mitochondrialcontrolregionandproteincodinggenesequencevariationamong phenotypicformsofbrowntrout Salmotrutta fromNorthernItaly. Mol.Ecol. 1994, 3,161–172.[CrossRef]
51. Patarnello,T.;Bargelloni,L.;Caldara,F.;Colombo,L.Cytochromeband16SrRNAsequencevariationinthe Salmotrutta (Salmonidae,Teleostei)speciescomplex. Mol.Phylogenet.Evol. 1994, 3,69–74.[CrossRef]
52. Gratton,P.;Allegrucci,G.;Gandolfi,A.;Sbordoni,V.Geneticdifferentiationandhybridizationintwonaturallyoccurring sympatrictrout Salmo spp.formsfromasmallkarsticlake. J.FishBiol. 2013, 82,637–657.[CrossRef]
53. Gratton,P.;Allegrucci,G.;Sbordoni,V.;Gandolfi,A.Theevolutionaryjigsawpuzzleofthesurvivingtrout(Salmotrutta L. complex)diversityintheItalianregion.AmultilocusBayesianapproach. Mol.Phylogenet.Evol. 2014, 79,292–304.[CrossRef]
54. Palombo,V.;DeZio,E.;Salvatore,G.;Esposito,S.;Iaffaldano,N.;D’Andrea,M.GenotypingoftwoMediterraneantrout populationsincentral-southernItalyforconservationpurposesusingarainbow-trout-derivedSNParray. Animals 2021, 11,1803. [CrossRef]
55. Karaman,S.PrilogpoznavanjuslatkovodnihribaJugoslavije.GlasnikSkopskognauˇcnogdruštvaknj. Skopje 1938, 18,131–139.
56. Bianco,P.G.AnupdateonthestatusofnativeandexoticfreshwaterfishesofItaly. J.Appl.Ichthyol. 2014, 30,62–77.[CrossRef]
57. Soldo,A.Firstmarinerecordofmarbletrout Salmomarmoratus J.FishBiol. 2013, 82,700–702.[CrossRef][PubMed]
58. Snoj,A.;Marˇceta,B.;Sušnik,S.;Melkiˇc,E.;Megliˇc,V.;Dovˇc,P.Thetaxonomicstatusofthe’seatrout’fromthenorthAdriaticSea, asrevealedbymitochondrialandnuclearDNAanalysis. J.Biogeogr. 2002, 29,1179–1185.[CrossRef]
59. Splendiani,A.;Ruggeri,P.;Giovannotti,M.;CaputoBarucchi,V.Roleofenvironmentalfactorsinthespreadofdomestictroutin Mediterraneanstreams. Freshwat.Biol. 2013, 58,2089–2101.[CrossRef]
60. Bianco,P.G.;Delmastro,G.B.Recentinovità tassonomicheriguardantiipescid’acquadolceautoctoniinItaliaedescrizionedi unanuovaspeciediluccio. Res.Wildl.Conserv. 2011, 2,1–14.
61. Gridelli,E. IPescid’AcquaDolcedellaVeneziaGiulia;DelBiancoD.:Udine,Italy,1935.
62. OldMapsOnline.Availableonline: https://www.oldmapsonline.org/ (accessedon18February2022).
63. Rossi,A.R.;Petrosino,G.;Milana,V.;Martinoli,M.;Rakaj,A.;Tancioni,L.Geneticidentificationofnativepopulationsof Mediterraneanbrowntrout Salmotrutta L.complex(Osteichthyes:Salmonidae)incentralItaly. Eur.Zool.J. 2019, 86,424–431. [CrossRef]
64. Zanetti,M.;NonnisMarzano,F.;Lorenzoni,M.ISalmonidiItaliani:LineeGuidaperlaConservazionedellaBiodiversità.2013, A.I.I.A.D.(AssociazioneItalianaIttiologiAcqueDolci)GruppodiLavoroSalmonidi.Availableonline: http://www.aiiad.it/sito/ temi/salmonidi/24-documento-salmonidi-febbraio-2013 (accessedon7February2022).
65. Lorenzoni,M.Thecheck-listoftheItalianfreshwaterfishfauna. Ital.J.Freshwat.Ichthyol. 2019, 5,239–254.
66. Pustovrh,G.;SušnikBajec,S.;Snoj,A.EvolutionaryrelationshipbetweenmarbletroutofthenorthernandthesouthernAdriatic basin. Mol.Phylogenet.Evol. 2011, 59,761–766.[CrossRef]
67. Pustovrh,G.;Snoj,A.;Bajec,S.S.Molecularphylogenyof Salmo ofthewesternBalkans,baseduponmultiplenuclearloci. Genet. Sel.Evol. 2014, 47,7.[CrossRef]
68. Lecaudey,L.A.;Schliewen,U.K.;Osinov,A.G.;Taylor,E.B.;Bernatchez,L.;Weiss,S.J.Inferringphylogeneticstructure,hybridizationanddivergencetimeswithinSalmoninae(Teleostei:Salmonidae)usingRAD-sequencing. Mol.Phylogenet.Evol. 2018, 124, 82–99.[CrossRef]
69. Bernatchez,L.;Guyomard,R.;Bonhomme,F.DNAsequencevariationofthemitochondrialcontrolregionamonggeographically andmorphologicallyremoteEuropeanbrowntrout Salmotrutta populations. Mol.Ecol. 1992, 1,161–173.[CrossRef]
70. Berrebi,B.;Povz,B.;Jesensek,D.;Cattaneo-Berrebi,G.;Crivelli,A.J.Thegeneticdiversityofnative,stockedandhybrid populationsofmarbletroutintheSocaRiver,Slovenia. Heredity 2000, 85,277–287.[CrossRef][PubMed]
71. Bernatchez,L.Theevolutionaryhistoryofbrowntrout(Salmotrutta L.)inferredfromphylogeographic,nestedclade,and mismatchanalysesofmitochondrialDNAvariation. Evolution 2001, 55,351–379.[CrossRef][PubMed]
72. Sanz,N.Phylogeographichistoryofbrowntrout:Areview.In BrownTrout:Biology,EcologyandManagement;Lobón-Cerviá,J., Sanz,N.,Eds.;Wiley&SonsLtd.:Hoboken,NJ,USA,2018;pp.17–64.
73. Maddison,W.P.Genetreesinspeciestrees. Syst.Biol. 1997, 46,523–536.[CrossRef]
74. SušnikBajec,S.;Pustovhr,G.;Jasenšek,D.;Snoj,A.PopulationgeneticSNPanalysisofmarbleandbrowntroutinahybridization zoneoftheAdriaticwatershedinSlovenia. Biol.Conserv. 2017, 184,239–250.[CrossRef]
75. Snoj,A.;Mari´c,S.;Berrebi,P.;Crivelli,A.J.;Shumka,S.;Sušnik,S.GeneticarchitectureoftroutfromAlbaniaasrevealedby mtDNAcontrolregionvariation. Genet.Sel.Evol. 2009, 41,22.[CrossRef]
76. Bianco,P.G.PotentialroleofthepaleohistoryoftheMediterraneanandParatethysbasinsontheearlydispersalofAuroMediterraneanfreshwaterfishes. Ichthyol.Explor.Freshwat. 1990, 1,167–184.
77. Meraner,A.;Baric,S.;Pelster,B.;DallaVia,J.Trout(Salmotrutta)mitochondrialDNApolymorphisminthecentreofthemarble troutdistributionarea. Hydrobiologia 2007, 579,337–349.[CrossRef]
78. Pujolar,J.M.;Lucarda,A.N.;Simonato,M.;Patarnello,T.Restrictedgeneflowatthemicro-andmacro-geographicalscalein marbletroutbasedonmtDNAandmicrosatellitepolymorphism. Front.Zool. 2011, 8,1–10.[CrossRef]
79. Lucarda,A.N.;Bargelloni,L.;Patarnello,T.;Gandolfi,G.Geneticcharacterisationof Salmotruttamarmoratus (Cuvier,1817) populationsbymeansofnuclearmarkers:Preliminaryresults. Quad.ETP 1999, 28,1–5.
80. Meraner,A.;Baric,S.;Pelster,B.;DallaVia,J.MicrosatelliteDNAdatapointtoextensivebutincompleteadmixtureinamarble andbrowntrouthybridizationzone. Conserv.Genet. 2010, 11,985–998.[CrossRef]
81. Marazzi,S. AtlanteOrograficodelleAlpi.SOIUSA:SuddivisioneOrograficaInternazionaleUnificatadelSistemaAlpino;Priuli&Verlucca: PavoneCanavese,Italy,2005.
82. Splendiani,A.;Ruggeri,P.;Giovannotti,M.;Pesaresi,S.;Occhipinti,G.;Fioravanti,T.;Lorenzoni,M.;NisiCerioni,P.;Caputo Barucchi,V.AlienbrowntroutinvasionoftheItalianpeninsula:Theroleofgeological,climateandanthropogenicfactors. Biol. Invasions 2016, 18,2029–2044.[CrossRef]
83. Splendiani,A.;Fioravanti,T.;Giovannotti,M.;Olivieri,L.;Ruggeri,P.;NisiCerioni,P.;Vanni,S.;Enrichetti,F.;CaputoBarucchi, V.Museumsamplescouldhelptoreconstructtheoriginaldistributionof Salmotrutta complexinItaly. J.FishBiol. 2017, 90, 2443–2451.[CrossRef][PubMed]
84. Casalis,G. DizionarioGeograficoStorico-Statistico-CommercialedegliStatidiS.M.ilRediSardegna;Maspero:Turin,Italy,1833; Volume1.
85. Casalis,G. DizionarioStorico-Statistico-CommercialedegliStatidiS.M.ilRediSardegna;Maspero:Turin,Italy,1852;Volume22.
86. Festa,E.IpescidelPiemonte. Boll.Mus.Zool.Anat.Comp.RUniv.Torino 1892, 7,1–125.
87. VonSiebold,C.T.E.UeberdieFischedesOber-Engadins.InProceedingsoftheVerhandlungenderSchweizerischenNaturforschendenGesellschaftzuSamaden,Samaden,Switzerland,24–26August1863;pp.173–190.
88. Monti,M. NotiziedeiPescidelleProvinciediComoeSondrioedelCantoneTicino;Franchi,C.:Como,Italy,1864.
89. Fatio,V. FaunedesVertébrésdelaSuisse:HistoireNaturelledesPoissons;IIpart;Georg,H.:Geneva,Switzerland;Basel,Switzerland, 1890;Volume5,pp.354–355.
90. Sommani,E.Il Salmomarmoratus CUV.:Suaorigineedistribuzionenell’Italiasettentrionale. Boll.PescaPiscic.EIdrobiol. 1960, 15, 40–47.
91. Schönswetter,P.;Stehlik,I.;Holderegger,R.;Tribsch,A.MolecularevidenceforglacialrefugiaofmountainplantsintheEuropean Alps. Mol.Ecol. 2005, 14,3547–3555.[CrossRef][PubMed]
92. Stefani,F.;Anzani,A.;Marieni,A.Echoesfromthepast:AgenetictraceofnativebrowntroutintheItalianAlps. Environ.Biol. Fish 2020, 102,1327–1335.[CrossRef]
93. Schorr,G.;Holstein,N.;Pearman,P.B.;Guisan,A.;Kadereit,J.W.Integratingspeciesdistributionmodels(SDMs)andphylogeographyfortwospeciesofAlpine Primula Ecol.Evol. 2012, 2,1260–1277.[CrossRef]
94. D’Ancona,I.J.;Merlo,S.LaspeciazionedelletroteitalianeedinparticolareinquelledellagodiGarda. Atti.Lst.Ven.Sci.Lett. Arti. 1959, 117,19–26.
95. Behnke,R.J.Thesystematicsofsalmonidfishesofrecentlyglaciatedlakes. J.FishRes.BoardCan. 1972, 29,639–671.[CrossRef]
96. Todesco,M.;Pascual,M.A.;Owens,G.L.;Ostevik,K.;Moyers,B.T.;Hubner,S.;Heredia,S.M.;Hahn,M.A.;Caseys,C.;Bock, D.G.;etal.Hybridizationandextinction. Evol.Appl. 2016, 9,892–908.[CrossRef]
97. Meldgaard,T.;Crivelli,A.J.;Jesensek,D.;Poizat,G.;Rubin,J.-F.;Berrebi,P.Hybridizationmechanismsbetweentheendangered marbletrout(Salmomarmoratus)andthebrowntrout(Salmotrutta)asrevealedbyin-streamexperiments. Biol.Conserv. 2007, 136, 602–611.[CrossRef]
98. Hamilton,K.E.;Ferguson,A.;Taggart,J.B.;Tómasson,T.;Walker,A.;Fahy,E.Post-glacialcolonizationofbrowntrout, Salmo trutta L.:Ldh-5*asaphylogeographicmarkerlocus. J.FishBiol. 1989, 35,651–664.[CrossRef]
99. Presa,P.;Krieg,F.;Estoup,A.;Guyomard,R.Diversité etgestiongénétiquedelatruitecommune:ApportdeI’étudedu polymorphismedeslocusprotéiquesetmicrosatellites. Gén.Sél.Evol. 1994, 26 (Suppl.S1),183s–202s.Availableonline: https://hal.archives-ouvertes.fr/hal-00894063 (accessedon20February2022).[CrossRef]
100. Largiadèr,C.R.;Scholl,A.EffectsofstockingonthegeneticdiversityofbrowntroutpopulationsoftheAdriaticandDanubian drainagesinSwitzerland. J.FishBiol. 1995, 47 (Suppl.A),209–255.[CrossRef]
101. Sommani,E.Sullapresenzadel Salmofario (L.)edel Salmomarmoratus (Cuv.)nell’Italiasettentrionale:Lorocaratteristiche ecologicheeconsiderazionirelativeairipopolamenti. Boll.PescaPiscic.Idrobiol. 1948, 3,136–145.
102. Keller,I.;Taverna,A.;Seehausen,O.EvidenceofneutralandadaptivegeneticdivergencebetweenEuropeantroutpopulations sampledalongaltitudinalgradients. Mol.Ecol. 2011, 20,1888–1904.[CrossRef]
103. Keller,I.;Schuler,J.;Bezault,E.;Seehausen,O.ParalleldivergentadaptationalongreplicatedaltitudinalgradientsinAlpinetrout. BMCEvol.Biol. 2012, 12,210.[CrossRef]
104. Analisidell’opera.NaturaMortaconPescidiEvaristoBaschenis.NaturaMortaconPesci,~1670,oilonCanvas, 65cm × 108cm. Bergamo,AcademyofFineArtsinCarrara.Availableonline: https://www.analisidellopera.it/natura-morta-con-pescibaschenis/ (accessedon22February2022).
105. Sacchi,B. (Platina)PlatinedeHonestaVoluptateetValetudine;deAquila,L.:Venice,Italy,1475.
106. Giovio,P. NovocomensisdePiscibusMarinis,Lacustribus,Fluviatilibus,ItemdeTestaceisacSalsamentisLiber;MinitiiCalvi,F.:Rome, Italy,1527.
107. Salviani,I. AquatiliumAnimaliumHistoriae,LiberPrimus:CumEorumdemFormis,AereExcusis;Saluianum:Rome,Italy,1554.
108. Porcacchi,T.;GiolitodeFerrari,G.;Gesuiti:Collegio,R. LaNobiltà dellaCittà diComoDescrittadaThomasoPorcacchidaCastiglione Arretino:ConlaTavoladelleCoseNotabili;diFerrariiGiolito,G.:Venice,Italy,1569.
109. Scappi,B. OperadiBartolomeoScappi,MastroDell’artedelCucinare,DivisainSeiLibri;de’Vecchi,A.:Venice,Italy,1570.
110. Grattarolo,B. HistoriadellaRivieradiSalò;Sabbio,V.:Brescia,Italy,1599.
111. Stefani,B. L’artediBenCucinare,etInstruireiMenPeritiinQuestaLodevoleProfessione:DoveAncos’InsegnaaFarPasticci,Sapori, Salse,Gelatine,Torte,etAltro;Osanna:Mantua,Italy,1662.
112. Roberti,G. LetteraSoprailCantode’Pesci;dellaVolpe,L.:Bologna,Italy,1767.
113. Delling,B.Morphologicaldistinctionofthemarbletrout, Salmomarmoratus,incomparisontomarbled Salmotrutta fromRiver Otra,Norway. Cybium 2002, 26,283–300.[CrossRef]
114. Snoj,A.;Glamuzina,B.;Razpet,A.;Zablocki,J.;Bogut,I.;Lerceteau-Köhler,E.;Pojski´c,N.;Sušnik,S.Resolvingtaxonomic uncertaintiesusingmolecularsystematics: Salmodentex andtheBalkantroutcommunity. Hydrobiologia 2010, 651,199–212. [CrossRef]
115. Pontalti,L.Latrotamarmoratadaifiumiairuscelli:Possibilità,perunaspecieinpericolo,diallungareilpropriohabitat.Seconda parte:Acclimatamentodiunapopolazione. Dendronatura 2020, 1,76–83.
116. Huitfeldt-Kaas,H. FerskvandsfiskenesUtbredelseogIndvandringiNorge;CentralTrykkeriet:Oslo,Norway,1918.
117. Sønstebø,J.H.;Borgstrøm,R.;Heun,M.Geneticstructureofbrowntrout(Salmotrutta L.)fromtheHardangerviddamountain plateau(Norway)analyzedbymicrosatelliteDNA:Abasisforconservationguidelines. Conserv.Genet. 2007, 8,33–44.[CrossRef]
118. Miró,A.;Ventura,M.EvidenceofexotictroutmediatedminnowinvasioninPyreneanhighmountainlakes. Biol.Invasions 2015, 17,791–803.[CrossRef]
119. Tiberti,R.;Splendiani,A.Managementofahighlyunlikelynativefish:Thecaseofarcticcharr Salvelinusalpinus fromtheSouthern Alps. Aquat.Conserv.Mar.Freshwat.Ecosyst. 2019, 29,312–320.[CrossRef]
120. Vilizzi,L.Thecommoncarp, Cyprinuscarpio,intheMediterraneanregion:Origin,distribution,economicbenefits,impactsand management. Fish.Manag.Ecol. 2011, 19,93–110.[CrossRef]
121. Merati,F.;Pascale,M.;Perosino,G.C.StoriadellatrotamediterraneainPiemonte. Cent.Ric.Ecol.Sci.Territ.(CREST).2021. Availableonline: https://green-crest.blog/area-tecnico-scientifica/ (accessedon22February2022).
122. ˇ Riˇcánková,V.P.;Robovský,J.;Riegert,J.;Zrzavý,J.RegionalpatternsofpostglacialchangesinthePalearcticmammaliandiversity indicateretreattoSiberiansteppesratherthanextinction. Sci.Rep. 2014, 5,12682.[CrossRef][PubMed]
123. Sabol,M.;Persico,D.;Troco,E.Firstfossilrecordofleopard-likefelid(Panthera cf. pardus)fromalluvialdepositsofthePoRiver innorthernItaly. Quat.Int. 2017, 436,192200.[CrossRef]
124. Salari,L.;Masseti,M.;Silvestri,L.LatePleistoceneandHolocenedistributionhistoryoftheEurasianbeaverinItaly. Mammalia 2019, 84,259–277.[CrossRef]
125. Anonymous.IlCastoroTornainAltoAdigeDopo400Anni.Trentino,2020.Availableonline: https://www.giornaletrentino.it/ cronaca/il-castoro-torna-in-alto-adige-dopo-400-anni-1.2467616 (accessedon6March2022).
126. EC—EuropeanCommission.ThePrecautionaryPrinciple:DecisionmakingunderUncertainty;FutureBrief2017,18.Produced fortheEuropeanCommissionDGEnvironmentbytheScienceCommunicationUnit,UWE,Bristol.Availableonline: http: //ec.europa.eu/science-environment-policy (accessedon13February2022).
127. Lucarda,A.N.;Patarnello,T.;Forneris,G.;Pascale,M.Populationgeneticsof S.truttamarmoratus, S.truttatrutta andtheirhybrid ofthePelliceRiverbasin(Piemonte,Italy). Quad.ETP 2000, 29,13–21.
128. McDonald,D.B.;Parchman,T.L.;Bower,M.R.;Hubert,W.A.;Rahel,F.J.Anintroducedandanativevertebratehybridizetoform ageneticbridgetoasecondnativespecies. Proc.Nat.Acad.Sci.USA 2008, 105,10837–10842.[CrossRef]
129. Aprahamian,M.W.;Smith,K.M.;McGinnity,P.;McKelvey,S.;Taylor,J.Restockingofsalmonids—Opportunitiesandlimitations. Fish.Res. 2003, 62,211–227.[CrossRef]
130. Arlinghaus,R.;Lorenzen,K.;Johnson,B.M.;Cooke,S.J.;Cowx,I.G.Managementoffreshwaterfisheries:Addressinghabitat, peopleandfishes.In FreshwaterFisheriesEcology;Craig,J.F.,Ed.;Wiley&Blackwell:Oxford,UK,2015;pp.557–579.[CrossRef]
131. Jellyman,P.G.;McHugh,P.A.;Simon,K.S.;Thompson,R.M.;McIntosh,A.R.Theeffectsofbrowntroutonthetrophicwebsof NewZealandstreams.In BrownTrout:Biology,EcologyandManagement;Lobón-Cerviá,J.,Sanz,N.,Eds.;Wiley:Hoboken,NJ, USA,2018;pp.569–598.
132. Schöffmann,J.;Mari´c,S.;Snoj,A.TroutofSoutheastEurope,WesternandCentralAsia.In TroutandCharoftheWorld;Kershner, J.L.,Williams,J.E.,Gresswell,R.E.,Lobón-Cerviá,J.,Eds.;AmericanFisheriesSociety:Bethesda,MD,USA,2019;pp.411–456.
133. Tiberti,R.;Brighenti,S.Doalpinemacroinvertebratesrecoverdifferentlyinlakesandriversafteralienfisheradication? Knowl. Manag.Aquat.Ecosyst. 2019, 420,37.[CrossRef]
134. Zarco,A.;Corbalán,V.;Debandi,G.PredationbyinvasiverainbowtroutonthecriticallyendangeredPehuenchespiny-chestfrog. J.FishBiol. 2020, 98,878–880.[CrossRef][PubMed]
135. Moritz,C.Defining‘evolutionarysignificantunits’forconservation. TrendsEcol.Evol. 1994, 9,373–375.[CrossRef]
136. Almodóvar,A.;Nicola,G.G.;Elvira,B.;García-Marín,J.L.IntrogressionvariabilityamongIberianbrowntroutevolutionary significantunits:Theinfluenceoflocalmanagementandenvironmentalfeatures. Freshwat.Biol. 2006, 51,1175–1187.[CrossRef]
137. Budy,P.;Rogers,K.B.;Kanno,Y.;Penaluna,B.E.;Hitt,N.P.;Thiede,G.P.;Dunham,J.;Mellison,C.;Somer,W.L.;DeRito,J. DistributionandstatusoftroutandcharinNorthAmerica.In TroutandCharoftheWorld;Kershner,J.L.,Williams,J.E.,Gresswell, R.E.,Lobón-Cerviá,J.,Eds.;AmericanFisheriesSociety:Bethesda,MD,USA,2019;pp.193–250.
138. Splendiani,A.;Fioravanti,T.;Giovannotti,M.;Negri,A.;Ruggeri,P.;Olivieri,L.;NisiCerioni,P.;Lorenzoni,M.;CaputoBarucchi, V.TheeffectsofpaleoclimaticeventsonMediterraneantrout:PreliminaryevidencesfromancientDNA. PLoSONE 2016, 11, e0157975.[CrossRef]
139. Kuwae,M.;Tamai,H.;Doi,H.;Sakata,M.K.;Minamoto,T.;Suzuki,Y.SedimentaryDNAtracksdecadal-centennialchangesin fishabundance. Commun.Biol. 2020, 3,558.[CrossRef]
140. Gary,M.K.;Nielsen,L.A.;Knight,R.L.;Schenborn,D.A. EcosystemManagement.Adaptive,Community-BasedConservation;Island Press:Washington,DC,USA,2002.