Spelljammer: Adventures in Space (D&D Campaign Collection - Adventure, Setting, Monster Book, Map, and DM Screen)
Wizards RPG Team Spelljammer: Adventures In Space (D&D Campaign Collection - Adventure https://ebookmass.com/product/spelljammer-adventures-in-space-ddcampaign-collection-adventure-setting-monster-book-map-and-dm-screenwizards-rpg-team-spelljammer-adventures-in-space-dd-campaigncollection-adventure/ ebookmass.com
Respiratory Biology of Animals
Respiratory Biology of Animals Evolutionary and Functional Morphology
Steven F. Perry
Institut für Zoologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Germany
Markus Lambertz
Institut für Zoologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Germany Sektion Herpetologie, Zoologisches Forschungsmuseum Alexander Koenig, Germany
Anke Schmitz
Institut für Zoologie, Rheinische Friedrich-Wilhelms-Universität Bonn, Germany
Great Clarendon Street, Oxford, OX2 6DP, United Kingdom
Oxford University Press is a department of the University of Oxford. It furthers the University’s objective of excellence in research, scholarship, and education by publishing worldwide. Oxford is a registered trade mark of Oxford University Press in the UK and in certain other countries
The moral rights of the authors have been asserted
First Edition published in 2019
Impression: 1
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, without the prior permission in writing of Oxford University Press, or as expressly permitted by law, by licence or under terms agreed with the appropriate reprographics rights organization. Enquiries concerning reproduction outside the scope of the above should be sent to the Rights Department, Oxford University Press, at the address above
You must not circulate this work in any other form and you must impose this same condition on any acquirer
Published in the United States of America by Oxford University Press 198 Madison Avenue, New York, NY 10016, United States of America
British Library Cataloguing in Publication Data
Data available
Library of Congress Control Number: 2019945437
ISBN 978–0–19–923846–0 (hbk.)
ISBN 978–0–19–923847–7 (pbk.)
DOI: 10.1093/oso/9780199238460.001.0001
Printed and bound by CPI Group (UK) Ltd, Croydon, CR0 4YY
We wish to dedicate this book to the memory of Steve Morris (1956–2009), whose untimely death was a great loss, not only for respiratory biology.
Preface
This book is meant to be a supplementary text for advanced courses in metabolic physiology, evolutionary biology, or related areas. As prerequisites we suggest courses in general biology, general zoology, animal physiology, and comparative anatomy. In addition, it may be of interest to students of medicine, biophysics, biochemistry, or related areas of molecular biology. To this end, we do not delve into the nuts and bolts of biochemistry, biophysics, and molecular biology, but rather emphasize the overarching evolutionary implications and basic principles that the study of respiratory biology can illustrate. But since special courses in respiratory physiology are not offered everywhere, we perhaps go into this area in a bit more detail than would be expected in a book on functional and evolutionary morphology. The same applies to the control of breathing, and some basic information must be supplied in order to understand the more detailed descriptions that follow. As we will discover again and again: ‘Before you can do what you want to do, you always have to do something else’. In general, it was our intention to provide an overview of the big picture of the evolution of respiration in animals. We consequently regularly cite
relatively recent overview or review articles on certain subjects rather than referencing all of the primary sources. This book was never intended to serve as an encyclopaedic reference volume. We therefore frequently selected interesting examples to illustrate a given phenomenon as a case in point, rather than providing a complete list of all known details in each and every species that has been examined. Furthermore and most importantly, you may not fnd all the answers you are interested in, but instead regularly will be confronted with persisting questions. This is in keeping with the primary intention of this book, which was to expose respiratory biology as a fertile ground for further research, in fact on all levels from molecular to evolutionary considerations. We hope that you will enjoy the read and in turn look forward to reading new advancements in this feld from your end as well, some of which may even be stimulated by critically reading the present volume.
Bonn, January 2019 The authors
Acknowledgements
First of all we would like to thank our publisher Oxford University Press for their incredible patience with us before this book eventually became fnished. We furthermore wish to express our sincerest thanks to those colleagues who read preliminary drafts of individual chapters and provided valuable criticism that helped us to improve the contents. These are, in alphabetical
order: Thorsten Burmester (Hamburg, Germany), Jon F. Harrison (Tempe, AZ, USA), John B. West (San Diego, CA, USA), Jürgen Markl (Mainz, Germany), Richard J.A. Wilson (Calgary, Canada). M.L. gratefully acknowledges the support of Michael H. Hofmann (Bonn, Germany) for providing the facilities and latitude for pursuing this and other projects.
‘The necessity of respiration to the support of life, and the evident injuries arising from any impediment in this function, induced the earliest medical philosophers to make it a subject of enquiry; and, from that time to this, it has afforded a continued subject of admiration, discussion, and dispute: not, indeed, of dispute, as whether it was necessary or not, but why it was necessary? what where the advantages the animal economy derived from thence? and by what means it was carried on?’
Robert Townson (1799). Tracts and Observations in Natural History and Physiology. London, printed for the author: p. 7.
CHAPTER 1
Prolegomena
Probably no process epitomizes life more than respiration. By respiration we mean the cascade of energy-producing biochemical reactions called oxidative phosphorylation, powered by a gradient of oxidation. If you are reading this book you probably already know that the terminal oxidant in most cases is atmospheric oxygen (O2), but a variety of micro-organisms also show respiration powered by other terminal molecules such as sulphur compounds or even carbon dioxide (CO2) or perchlorate (e.g. Liebensteiner et al., 2016 and references therein). The end products of respiration are phosphorylated nucleotides such as adenosine triphosphate (ATP) or guanosine triphosphate (GTP) and, as waste products, carbon dioxide and water. As a side effect of respiration, many of the reactions involved in oxidative phosphorylation are exothermic. The heat released can aid in the maintenance of a high, constant body temperature (homeothermy), but in extremely large animals such as sauropod dinosaurs the sheer amount of heat produced may even result in substantial physiological problems (gigantothermy).
However, the ‘cellular respiration’ just described represents only the fnal phase (here called phase 5). In many organisms, respiration also includes four other phases, which, now working back from the cells to the outside world are (4) gas consumption and production at the tissue and cellular level, (3) the transport of these gases between the gas exchanger and tissues via body fuids (e.g. a circulatory system, if present), (2) diffusive gas exchange between the medium (air or water) and the organism, and fnally (1) the convection of oxygen and carbon
dioxide to and from a gas exchanger (e.g. lung, gill, skin, or tracheae). Phase 4 is also referred to as ‘aerobic metabolism’, phase 3 as ‘circulation’ or ‘perfusion’, phase 2 is ‘gas exchange’, and phase 1 is ‘breathing’ or ‘ventilation’.
The focus of this book is the evolution of respiratory systems in animals. But in order to really understand this, we must frst deal with some basic concepts of respiration, including the properties of the gases involved and the historical sequence of events that led to the development of respiratory biology as we know it today. We then move on to some basics of functional morphology and physiology as they relate to respiration. But no organ system can be studied on its own, since all of them interact with each other to comprise an organism. So, to be more precise, we will deal in detail with the evolution of the structure–function interaction in respiratory physiology and anatomy. The term ‘faculty’ as defned by Bock and von Wahlert (1965) describes exactly this structure–function interaction, whereby every structure can have multiple functions and every function can involve numerous structures.
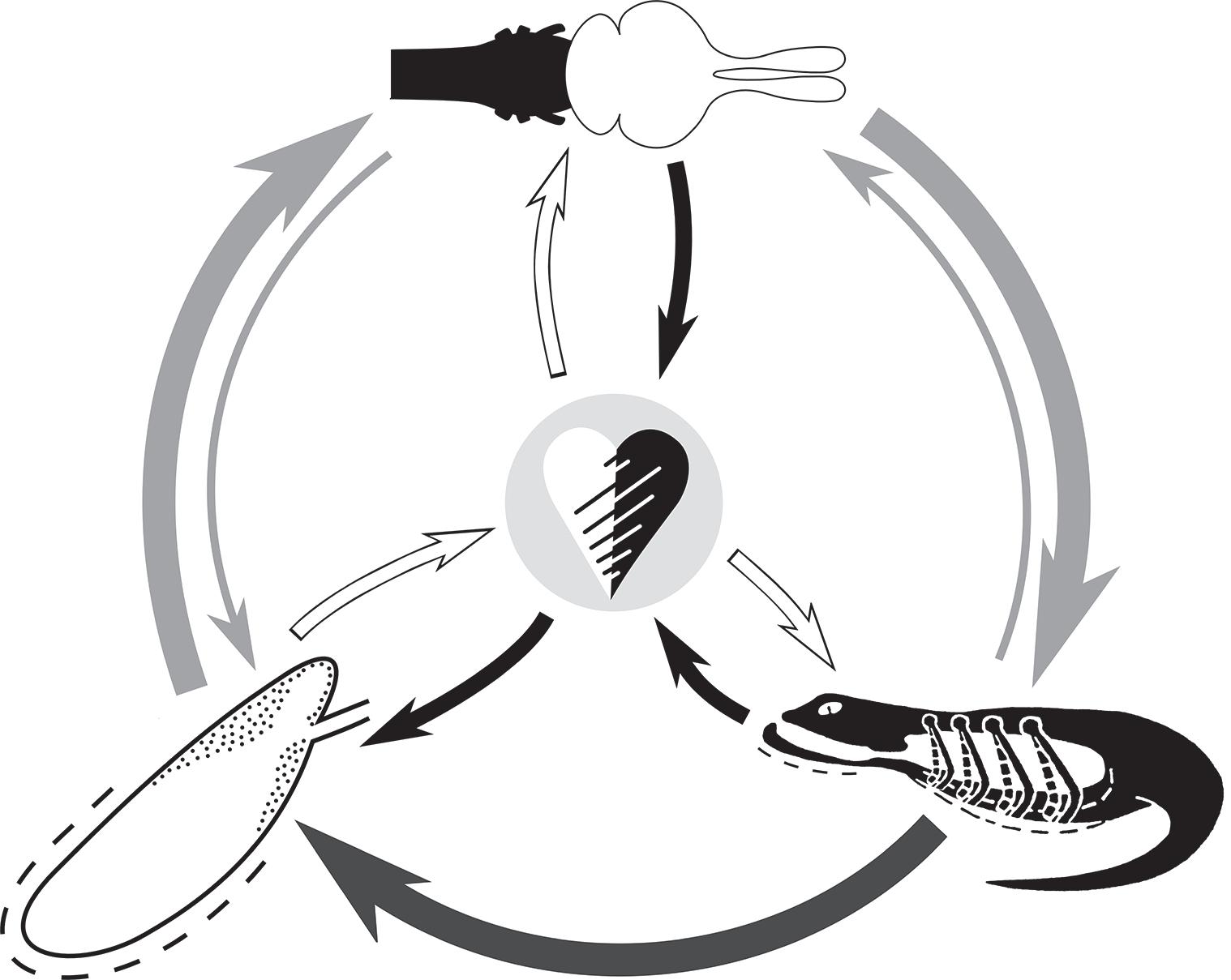
The ‘faculty’ concept is based on the assumption that a given function—in this case respiration—is related to a variety of anatomical components, thereby composing a functional unit (Figure 1.1). This idea is not new and was probably best expressed by the French comparative and functional morphologist and founder of reconstructive palaeontology, Georges Cuvier (1769–1832), in his so-called law of the correlation of the parts (Cuvier, 1812). There he
Figure 1.1 A schematic representation of the concept of a respiratory faculty, exemplifed in a lizard. Brainstem shown in black. Active respiratory pump illustrating only costal breathing and a simplifed rib cage. Gray arrows indicate direct biomechanical (dark shading) and neuronal (light shading) connections, whereby the thinner arrows indicate feedback. Black or white arrows indicate an indirect loop via the circulatory sytem, here with an intercardiac shunt possibility: Black arrows and print, deoxygenated blood; white arrows and print, oxygenated blood. The size and colour of the print (O2 or CO2) indicates the predominance of this gas in the circulatory loop. Original M.L. and S.F.P in Hsia et al. (2013).
states that an animal is a closed unit in which all component parts mutually correspond and unite in reciprocal reaction, such that no one part can be changed without changing all of the others. Focusing on the cardiorespiratory faculty, Weibel and Taylor (1981) coined the term ‘symmorphosis’. In a series of publications over at least the next 10 years (see Weibel et al., 1991) they applied physiological and quantitative morphological methods to determine just how close structure and function are coupled with regard to oxygen supply and demand in mammals. Although within closely related phylogenetic groups symmorphosis appeared to generally apply, detailed independent statistical analysis revealed weaknesses in the general validity of this principle (Garland and Huey, 1987). The latter authors concluded: ‘Although symmorphosis has heuristic value as a working hypothesis, it should not presently be considered an established principle; available data appear largely contradictory’. It is just this heuristic value that causes symmorphosis to surface as a hypothesis even in recent publications (Gifford et al., 2016).
As you read along, you will notice two overriding principles that appear and reappear at all organizational levels from molecular to organismic. The frst of these can be summarized as ‘Getting it right the frst time’. Basically, you can’t kill a good idea! A couple of examples illustrate how this works. When we look more closely at the evolution of the respiratory faculty it will become evident that sometimes functions change while structures are conserved. At the molecular level, for example, we see nucleotides such as adenosine appearing with multiple functions ranging from a building block of nucleic acids to energy transport, and they can even assume new functions, such as infuencing oxygen affnity of oxygen-carrying molecules such as haemoglobin. An example at the organ level is the lung. This primary gas-exchange organ also assumes crucial physiological functions such as the conversion of angiotensin I to angiotensin II, or as a site of detoxifcation and immune activity. But sometimes function is conserved or improved upon while the structures around it radically change. This can be recognized at any organizational level and is not limited to respiratory biology. As an example at the cellular physiological level, we see a plethora of socalled respiratory proteins that serve in oxygen transport. At the organ level we fnd, for example, countercurrent gas exchange being conserved in fsh gills in spite of radically different ventilatory mechanisms and gill structures in hagfsh, lampreys, sharks, and bony fsh. And sometimes a primary function becomes secondary, while an ancestral secondary function becomes primary. An example for such an exaptation at the organ level is the pseudobranch in fshes. What used to be a gas-exchange organ (gill) that contained oxygen receptors, lost its primary gas-exchange function but retained the oxygen receptors as an important monitoring organ for oxygen tension in the blood.
The second reoccurring principle is the evolutionary cascade. This positive feedback phenomenon dovetails in an evolutionary context with exaptation when the exaptive faculty complements with others synergistically. One example is the avian respiratory faculty. The essential structural elements were probably already present in non-avian dinosaurs or even in ancestral archosaurs, but it could only become exploited to its limits in actively fying
birds, an evolutionary quirk that could not have been predicted.
Evolution leaves tracks. This will become evident in every chapter of this book. But certainly the most far-reaching of these tracks is the realization that each plant and animal, including us humans, carries within every cell a molecular memory of the conditions under which life originated and which are still necessary. To understand what we are talking about, let us journey back to a time about 4–3.5 thousand million years ago (mya). We must imagine a hot atmosphere consisting mostly of water vapour and gases such as nitrogen (N2), carbon dioxide, hydrogen sulphide, sulphur dioxide, and traces of ammonia and methane, but no free oxygen. Sort of like living in the mouth of a volcano. The world was spinning much faster than today, meaning that the days and nights were much shorter than they are now. But more importantly, this must have resulted in tremendous winds. As the Earth cooled, water vapour condensed to water and the seas formed. The Moon was much closer to the Earth than it is now, resulting in gigantic tides. As the fog dissipated to form clouds, the Sun’s rays could reach the surface unfltered by an ozone (O3) layer, since oxygen was lacking in the atmosphere. The sunlight contained high-energy ultraviolet rays that would certainly have been lethal to any life form exposed to them. It is diffcult to imagine the origin of life under these violent conditions.
A recent scenario for the origin of life suggests that it all began in the deep sea, near sulphur vents (Lane and Martin, 2012). This hypothesis holds that life began from organic macromolecules reacting with each other in a closed, sulphur-rich environment that was relatively stable compared with what was going on at the surface. We speculate that this primordial life was repeatedly brought to the surface by upwelling activity and washed into inland seas by the immense tides. Under these conditions, a sort of ‘training’ could have taken place by which proto-life forms were exposed under reasonably calm conditions to relatively stable molecular oxygen and highly unstable oxygen and hydroxyl radicals, generated during photolysis of the water by the ionizing radiation. The life forms that were able to utilize oxygen rather than sulphur as a terminal oxidant would have had a great advantage over those which
were not, simply due to the immense surplus of oxygen at the surface relative to sulphur. In other words, we envision a two-step origin of life: the primary origin of cellular organisms in the deep sea and the modifcation of these organisms for survival through the development of circadian gene activity programmes and the ability to detoxify or utilize oxygen and oxygen radicals.
Rainfall and flling of the inland seas together with the high tides could have brought life back to the sea. This means that the capability to deal with oxygen in a way that is not only compatible with but also advantageous to life could have originated long before the photosynthetic origin of atmospheric oxygen (see later in this chapter) and served both inland sea and marine organisms. In the course of this book you will discover the tracks of many of these stages conserved within the evolutionary history of respiration in animals. Then something happened.
About 2 thousand mya, the most complex forms of life were bacteria: among them, photosynthetic cyanobacteria. They were the cutting edge of life back then since they could use the sun’s energy to power the assimilation of carbon dioxide and its conversion into metabolic intermediates and structural macromolecules. This process poured out molecular oxygen, which began to accumulate in the atmosphere. By somewhere between 900 and 600 mya it reached about 1 per cent of the present level. This is called the ‘Pasteur point of evolution’, since it was the famous French chemist and microbiologist Louis Pasteur (1822–1895) who discovered that oxygen paradoxically decreases the frst steps of metabolism while at the same time stimulating growth of yeast cells in culture. But this low level of atmospheric oxygen was still suffcient for many organisms to switch to the highly effcient aerobic form of metabolism. As we shall see later, it is just this level of oxygen that is maintained in our tissues and cells to this day: we are carrying with us an ancient environment dating back to the early days of aerobic life.
Among the ancient eubacteria were also ones that had been internalized by archaea. These endosymbionts could utilize not only molecular oxygen but also acetyl coenzyme A: an adenosine-based substance produced by the host. These endosymbiotic bacteria, now called mitochondria, later became assimilated by nucleated cells (eukaryotes) and gave
them the ability to produce about 16 times as much ATP from each mole of glucose as could those that didn’t have mitochondria. ATP is produced by phosphorylation of adenosine diphosphate (ADP) either anaerobically by glycolysis in the cytosol or aerobically in the mitochondria. Although cyanobacteria initially changed the atmosphere to an oxidizing one, the large amounts of atmospheric oxygen—reaching peak levels of nearly double present-day levels—were later the result of photosynthesis of green plants, which through independent endosymbiotic processes had acquired chloroplasts.
Mitochondria produce ATP as we have already seen, but equally importantly, they remove oxygen from the cytosol and keep the level similar to that prevailing when oxygen began to accumulate in the atmosphere. This is important, because more oxygen results in higher levels of ATP, which in turn puts the brakes on metabolism right at the glycolytic level (Pasteur effect). So these aerobic eukaryotes were exapted for high energy production in a high-oxygen atmosphere that would come later, by keeping the oxygen in the cells at a low, ‘ancient’ level!
Later, the actual timing is still highly debated, multicellular eukaryotic organisms evolved and what we nowadays call animals (Metazoa) entered the game. And this, in a nutshell, takes us up to the point where the story of the structure, function, and evolution of respiratory faculties in major animal groups begins. So join us on this journey!
We shall be travelling on the evolutionary biology ship, so before embarking we need to have a look at some of the guidelines for a safe and unencumbered journey. First of all we need to use a vocabulary that may not be familiar to everyone and may even appear cumbersome. For example, nothing ever evolves ‘for’ a specifc purpose, so even implications of such teleological thinking are categorically avoided. Having said this, we realize that epigenetics does exist and that many adaptive phenomena appear to conform
to teleological guidelines. But even adaptability is genetically anchored and does not escape evolutionary selection.
Another minefeld that needs to be briefy touched on here is the species problem. The specifc identity of the organisms in question remains uncertain in many cases. We thus frequently also just refer to a certain genus instead of a specifc species, well aware that the details in fact may differ between congeneric species. We refer to this as the ‘taxonomic uncertainty relation’. The examples provided within the present book are meant to illustrate the diversity of the respiratory faculty exhibited among taxa. In other words, we focus on the big picture rather than being comprehensive with regard to every species-level specialization.
This leads us to the topic of ranks. Many textbooks still use ‘phyla’ or ‘orders’ for groupings such as craniotes or turtles, respectively. We completely avoid referring to ranks and adhere to a more phylogenetic framework, which also makes it inappropriate to speak of ‘primitive’ species or taxa. For example, hagfsh are not primitive craniotes, but a basally branching radiation of them. Speaking of craniotes: most commonly this group colloquially is called vertebrates, but we prefer the name Craniota and its derivative craniotes (Lambertz, 2016a). Similarly with hexapods, traditionally called insects, which based on our current understanding are only a subordinated taxon of the Hexapoda. However, you will fnd the term insects along with hexapods within this book, because most research has been done on ‘proper’ insects. Having said all of this, we occasionally also deviate from a strictly phylogenetic framework and adopt a more grade-like than cladelike approach if summarizing certain general phenomena, observable in, for instance, ‘reptiles’ rather than speaking of non-avian and non-mammalian amniotes. The same applies to the usage ‘invertebrates’ as opposed to non-craniote animals.
So that’s it. Let’s see where this trip takes us.
2
A very brief history of respiratory biology
A better understanding of what life is and how living organisms function has always been of crucial importance to humans, but ‘biology’ as a scientifc discipline is quite young, the term being coined around 1800. Similarly, ‘respiratory biology’ as a discrete branch of biology is much younger and even today the term is not commonly used. However, the study of respiration-relevant processes and phenomena does look back on a long history. In the course of this book we will often give the names of scientists together with their dates of birth and death when their crucial fndings are mentioned for the frst time. The purpose of the present brief chapter is to place many of the major advances in respiratory biology and the names of key scientists in historical perspective and to point out their importance to the development of what we now call respiratory biology. Clearly this chapter—just like the rest of the book—refects to some extent the authors’ own interests, and we do not intend a comprehensive and unbiased approach. For this the reader is encouraged to consult other excellent works such as those of Fishman and Richards (1982), Otis (1986), SchmidtNielsen (1995), West (1996, 2011, 2012, 2015, 2016), Gunga (2009), and Fitzgerald and Cherniack (2012) and the references found there.
The clinical importance of lungs and breathing is recorded in papyrus documents dating back thousands of years, and Aristotle (384–322 bc) postulated vital power in the redness of blood. But the frst suggestion that a vital substance is actually in the
air taken up in the lungs and transported by blood vessels to the heart and from there to the rest of the body was made by Galen (c.129–200 or 216), gladiator surgeon and private physician to the Roman emperor Marcus Aurelius (121–180). The knowledge about structure and function of the human body in general and respiration in particular was codifed in the works of the authorities Aristotle and Galen, and new investigation was strongly discouraged in the Christian world, until it surfaced during the Italian Renaissance. In Padua, the Spanish-born surgeon and theologian Miguel Serveto (1509 or 1511–1553), co-founder of Unitarianism, was the frst to postulate pulmonary circulation (interestingly, published within his theological opus magnum of 1553, for which he was burned at the stake!). The frst demonstration of capillaries in general, however, was provided much earlier by none other than Leonardo da Vinci (1452–1519) using wax injections of human cadavers, but it was never published. Among the frst to describe the comparative anatomy of the craniote respiratory system was Claude Perrault (1613–1688). In 1669, he published illustrations of some 90 dissections, including chameleons with their aberrant lungs. G. Joseph Duverney (1648–1730) continued this work after Perrault’s death.
Meanwhile, in the area of cardiorespiratory physiology, the prediction of blood circulation in 1628 by the English surgeon and comparative physiologist William Harvey (1578–1657) caused quite a stir, because it refuted both the Aristotelian concept that
blood circulated due to its own vital power and Galen’s belief that vitalized, nutrient-rich blood was consumed by body tissues and not returned to the heart. In addition, the idea that the heart was just a pump rather than a refection of soul and mood just did not sit well. Harvey, a student of Hieronymus Fabricius ab Acquapendente (1533/37–1619), worked in Padua as well, but it was not until in 1661 in Bologna that Marcello Malpighi (1628–1694) actually observed blood entering and leaving a microvascular bed in the frog lung. Unaware of Malpighi’s work, Antonie van Leeuwenhoek (1632–1723) made similar observations and another Dutch scientist, Jan Swammerdam (1637–1680), also made substantial pioneering contributions to pulmonary function. This, together with biomechanical studies of breathing based on the mechanistic philosophical tradition dating back to René Descartes (1596–1650), made possible a purely biophysical approach to respiration and opened the door to mathematical modelling or simulation of gas exchange and breathing mechanics by subsequent generations of researchers. As the pieces were coming together regarding the structure and function of the animal body, including its respiratory faculty, the magic also was slowly disappearing from alchemy. It was known that something required for life was present in air, and for lack of a better word, it was called ‘phlogiston’. Comparisons were made between the burning of a candle and respiration, both of which consumed ‘phlogistonated’ air and resulted in ‘fxed’ or ‘dephlogistonated’ air and water. Although the English theologian and chemist Joseph Priestley (1733–1804) is usually credited with the discovery of oxygen, the French chemist Antoine Laurent de Lavoisier (1743–1794) and the Swedish chemist Carl Wilhelm Scheele (1742–1786) may also share this honour. Heating mercuric oxide had been long known to alchemists as a way of producing quicksilver, but the gas released had been ignored. Now that oxygen had been identifed it could be used in experiments. The Russian chemist Dmitri Mendeleev (1834–1907) is usually credited with the development of the periodic table in 1869. However, numerical constants corresponding to atomic weights were known to the British eccentric chemist Henry Cavendish (1731–1810) a century earlier. By careful measurement of weights and volumes, Cavendish
discovered that water is the oxide of hydrogen, the gas he is credited with discovering, although he expressed the reaction in terms of phlogiston. So by the end of the eighteenth century it was known that aerobic metabolism was chemically comparable to combustion. On this basis, in 1800 the Cornish chemist, Sir Humphrey Davy (1778–1829), in addition to conducting numerous other experiments that predated modern biochemistry, exposed arterial and venous blood to a vacuum and demonstrated that oxygen and carbon dioxide, respectively, were transported, but it still was a long way to our current understanding of the respiratory process.
In the footsteps of the Swiss Albrecht von Haller (1708–1777), the German Johann Friedrich Blumenbach (1752–1840) was among the leading anatomists and physiologists during the second half of the eighteenth century. In 1787, with the intention of explaining the different kinds of movements in animals, he postulated a vita propria, a special or inner life, for the movements of those organs and structures that he was otherwise unable to explain: not unlike Aristotle’s vitalist views on the circulation of blood. Vita propria was assumed to account for ventilation of the lungs (i.e. spontaneous movement of the lungs was thought to move the ribs and not the reverse). It was actually one of Blumenbach’s rather lesser-known students who frst challenged this view: the English-born Robert Townson (1762–1827). He re-embarked on the path of Malpighi, Swammerdam, and others and conducted experiments on the respiratory movements in amphibians and turtles during the 1790s. He completely rejected the idea of the vita propria and instead declared movements of the throat and those of the lateral fanks, respectively, as responsible for ventilation in these animals (Figure 2.1). As we shall see later on, it was not until the second half of the twentieth century and the use of sophisticated electromyographic recordings that his pioneering work eventually became corroborated and accepted.
The nineteenth century could be considered the golden age of comparative anatomy and embryology. Beginning with the works of the French Georges Cuvier, the German Johann Friedrich Meckel the younger (1781–1833), and the German Martin Heinrich Rathke (1793–1860), accurate descriptions of the developing and mature respiratory organs of