Edited by Jennifer A. Dunne, H. Charles J. Godfray, and Ben Sheldon
The Comparative Method in Evolutionary Biology
Paul H. Harvey and Mark D. Pagel
The Cause of Molecular Evolution
John H. Gillespie
Dunnock Behaviour and Social Evolution
N. B. Davies
Natural Selection: Domains, Levels, and Challenges
George C. Williams
Behaviour and Social Evolution of Wasps: The Communal Aggregation Hypothesis
Yosiaki Ito
Life History Invariants: Some Explorations of Symmetry in Evolutionary Ecology
Eric L. Charnov
Quantitative Ecology and the Brown Trout
J. M. Elliott
Sexual Selection and the Barn Swallow
Anders Pape Møller
Ecology and Evolution in Anoxic Worlds
Tom Fenchel and Bland J. Finlay
Anolis Lizards of the Caribbean: Ecology, Evolution, and Plate Tectonics
Jonathan Roughgarden
From Individual Behaviour to Population Ecology
William J. Sutherland
Evolution of Social Insect Colonies: Sex Allocation and Kin Selection
Ross H. Crozier and Pekka Pamilo
Biological Invasions: Theory and Practice
Nanako Shigesada and Kohkichi Kawasaki
Cooperation Among Animals: An Evolutionary Perspective
Lee Alan Dugatkin
Natural Hybridization and Evolution
Michael L. Arnold
The Evolution of Sibling Rivalry
Douglas W. Mock and Geoffrey A. Parker
Asymmetry, Developmental Stability, and Evolution
Anders Pape Møller and John P. Swaddle
Metapopulation Ecology
Ilkka Hanski
Dynamic State Variable Models in Ecology: Methods and Applications
Colin W. Clark and Marc Mangel
The Origin, Expansion, and Demise of Plant Species
Donald A. Levin
The Spatial and Temporal Dynamics of Host–Parasitoid Interactions
Michael P. Hassell
The Ecology of Adaptive Radiation
Dolph Schluter
Parasites and the Behavior of Animals
Janice Moore
Evolutionary Ecology of Birds
Peter Bennett and Ian Owens
The Role of Chromosomal Change in Plant Evolution
Donald A. Levin
Living in Groups
Jens Krause and Graeme D. Ruxton
Stochastic Population Dynamics in Ecology and Conservation
Russell Lande, Steiner Engen, and Bernt-Erik Sæther
The Structure and Dynamics of Geographic Ranges
Kevin J. Gaston
Animal Signals
John Maynard Smith and David Harper
Evolutionary Ecology: The Trinidadian Guppy
Anne E. Magurran
Infectious Diseases in Primates: Behavior, Ecology, and Evolution
Charles L. Nunn and Sonia Altizer
Computational Molecular Evolution
Ziheng Yang
The Evolution and Emergence of RNA Viruses
Edward C. Holmes
Aboveground–Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change
Richard D. Bardgett and David A. Wardle
Principles of Social Evolution
Andrew F. G. Bourke
Maximum Entropy and Ecology: A Theory of Abundance, Distribution, and Energetics
John Harte
Ecological Speciation
Patrik Nosil
Energetic Food Webs: An Analysis of Real and Model Ecosystems
John C. Moore and Peter C. de Ruiter
Evolutionary Biomechanics: Selection, Phylogeny, and Constraint
Graham K. Taylor and Adrian L. R. Thomas
Quantitative Ecology and Evolutionary Biology: Integrating Models with Data
Otso Ovaskainen, Henrik de Knegt, and Maria del Mar Delgado
Mitonuclear Ecology
Geoffrey E. Hill
Mitonuclear Ecology
GEOFFREY E. HILL
Professor, Department of Biological Sciences, Auburn University, USA
Great Clarendon Street, Oxford, OX2 6DP, United Kingdom
Oxford University Press is a department of the University of Oxford. It furthers the University’s objective of excellence in research, scholarship, and education by publishing worldwide. Oxford is a registered trade mark of Oxford University Press in the UK and in certain other countries
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, without the prior permission in writing of Oxford University Press, or as expressly permitted by law, by licence or under terms agreed with the appropriate reprographics rights organization. Enquiries concerning reproduction outside the scope of the above should be sent to the Rights Department, Oxford University Press, at the address above
You must not circulate this work in any other form and you must impose this same condition on any acquirer
Published in the United States of America by Oxford University Press 198 Madison Avenue, New York, NY 10016, United States of America
British Library Cataloguing in Publication Data
Data available
Library of Congress Control Number: 2018964761
ISBN 978–0–19–881825–0 (hbk.)
ISBN 978–0–19–881826–7 (pbk.)
DOI: 10.1093/oso/9780198818250.001.0001
Printed in Great Britain by Bell & Bain Ltd., Glasgow
Links to third party websites are provided by Oxford in good faith and for information only. Oxford disclaims any responsibility for the materials contained in any third party website referenced in this work.
Preface
Birdwatching brought me to mitonuclear ecology. As a pre-teen, I was introduced to birding by a high school biology teacher, and I quickly became obsessed with seeing all the birds in my Golden Field Guide to the Birds of North America. I certainly didn’t appreciate it at the time, but a field guide to the birds of a region, through beautiful color illustrations, presents some of the most significant unexplained patterns in the natural world. All birds are neatly binned into species; the field guide suggests no ambiguity. Each species has a discrete range. Some birds thrive exclusively in warm climates. Other birds that seem identical in size, shape, and life history thrive in cold climates. Some birds are colorful; some are not. Like people, all birds come in two sexes. In some birds, the sexes look alike; in others, they do not. Explaining these patterns so starkly revealed in the pages of a field guide became the academic passion of my life. But a decade into the new millennium, despite 30 years of effort, I felt that I had made frustratingly little progress toward answering any of the basic questions that continued to confront me each time I opened a field guide. Then, I read Power, Sex, Suicide: Mitochondria and the Meaning of Life by Nick Lane, and for the first time I started to think about the major patterns of bird diversity from the perspective of the evolution of eukaryotes and the coevolution and coadaptation of mitochondrial (mt) and nuclear (N) genomes. It was as if I had turned on a light in a dark room.
In this book, I summarize emerging new theory and empirical observations that reinterpret key features of eukaryotic life in light of the necessity of coadapation of co-functioning mt and N genes. I call this emerging line of research mitonuclear ecology. It is the integration of studies of the coevolution and coadaptation of mt and N gene products into investigations of the evolution and ecology of whole organisms in natural environments. My purpose in creating this book is to present the new ideas and empirical observations that underlie mitonuclear ecology as an important new research focus. I propose that evolutionary ecology is poised to make a substantial leap forward in understanding fundamental features of complex organisms as evolutionary ecologists fully consider the implications of coadaptation and coevolution of mt and N genes to enable core life processes.
The origins of mitonuclear ecology lay in the biochemistry and cell biology laboratories that, in the mid-twentieth century, were in a different universe than lab groups focused on the evolution of whole organisms. In 1961, Peter Mitchell made one of the great intellectual leaps in the history of science when he proposed that chemiosmosis was the mechanism for oxidative phosphorylation and aerobic respiration in the mitochondria of eukaryotes (Mitchell, 1961) (Box 1.1). Mitchell’s brilliant discovery
was made at about the same time as the discovery that the mitochondrion carried its own genome (Nass and Nass, 1963; Schatz et al., 1964). Within a few years following these breakthroughs, cell biologists deduced that some components of the electron transport system were encoded by mt genes, while other components were encoded by N genes (Borst and Grivell, 1973). These were monumental discoveries by reductionistic biologists, but they went essentially unnoticed by organismal biologists focused on understanding evolution of plants and animals in natural environments. As a graduate student in the evolutionary biology program at the University of Michigan in the 1980s, I was taught nothing about oxidative phosphorylation, chemiosmosis, Peter Mitchell, or the function of mt genes. I was informed that mt genes were housekeeping genes, that variation in mt genotype was functionally neutral, and that the only reason to pay attention to mt DNA was for phylogenetic reconstructions. In retrospect, it was the biggest failure of my education in biology.
In the spring of 2012, I saw a reference to the book Power, Sex, Suicide: Mitochondria and the Meaning of Life by Nick Lane (2005) and on a whim I ordered a copy. I had never heard of Lane, and the topic of the book—mitochondria and the origins of complex life—was far from my focal interests as a behavioral ecologist studying plumage coloration in birds (Hill, 2002). By the end of the first chapter, I was captivated. It is not an exaggeration to say that Lane’s book fundamentally changed my academic life and the direction of my career as a biologist. What I found in Lane’s book was a narrative that shook the foundation of my view of biology—at once showing me the relevance of subcellular biology and forcing me to rethink fundamental aspects of evolutionary theory. I shared the book with my colleague, Jim Johnson, who is a biochemist well acquainted with mitochondrial processes, and Jim too was captivated. Jim was reading ahead of me and he sent me a note suggesting that I should page ahead to the section on the genes that code for the electron transport system. Jim wrote to me: “Lane makes the argument that the compatibility between the DNA in the mitochondria . . . and the nucleus is critical for overall respiratory efficiency.” Jim’s note was my first introduction to the concept of mitonuclear compatibility. Jim and I were already thinking and writing about the role of mitochondrial function in the production of ornamental red pigments in birds (Hill and Johnson, 2012; see also Chapter 7). After reading Lane’s book we began to contemplate the role of mitonuclear interaction in condition-dependent sexual signaling. Soon thereafter, we set to work on outlining a theory of sexual selection whereby ornaments signal mitonuclear compatibility (Hill and Johnson, 2013).
Conceptualizing how mates could be sorted for mitonuclear compatibility by assessment of ornaments was one of the greatest intellectual challenges of my career, and, in considering how mate choice might maintain mitonuclear coadaptation, I came to realize that mitonuclear interactions lie at the heart of not only sexual selection but also the process of speciation and the nature of species. The role of mitonuclear coevolution in the process of adaptation fell into place and soon I was viewing essentially all of the major features of complex life through mitonuclear lenses. This book is an attempt to organize new thinking (emerging in my own head as well as within the subdiscipline of evolutionary ecology) about how mitonuclear interactions shape complex life.
The theme of this book is mitonuclear coadaptation and coevolution and explicitly not broader cytonulear interactions. I scarcely mention plastid genomes in these chapters. I think there can be merit to making theoretical arguments as universal as possible (and hence I resisted my inclination to focus this book only on animal mitonuclear ecology), but there comes a point at which too broad of a focus is no focus at all. It will be challenging enough for field ecologists to wade through my opening chapters focused on genomic architecture, cell biology, and respiratory pathways. It would be counter-productive to follow these chapters with additional chapters on photosynthesis and a second set of genomic interactions between chloroplast and N genomes. Mitonuclear interactions are the common threads that bind all eukaryotes. There are, to be sure, many parallels in mitonuclear and chloronuclear genomic interactions, but I leave the development of plastid nuclear ecology for other evolutionary biologists.
I make no attempt to be balanced in my presentation of ideas and hypotheses. The purpose of this book is to succinctly outline the basic principles of mitonuclear ecology and to advocate for the importance of this approach to evolutionary questions. Topics such as the evolution of sex, the nature of species, and the process of sexual selection all have histories of investigations stretching back decades and a literature of hundreds if not thousands of papers. A balanced and comprehensive review of any of these topics would be a book-length project in itself, and such reviews are already available. I draw on previous literature primarily to set the stage for the new hypotheses emerging from consideration of the coadaptation of mt and N genomes.
My target audience is organismal biologists with limited training in cellular respiration and cell biology. With an audience of biologists, I don’t take the space to explain basic concepts like transcription, translation, natural selection, genetic drift, and so forth, but I do try to be careful in what I take for granted regarding previous knowledge of biochemistry and cell biology. It is my hope that not only organismal biologist interested in cellular-level processes but also cell biologists interested in macro-evolution will find this book interesting. It is in the integration of genomics and biochemistry with ecology and organismal biology that the power of mitonuclear ecology lies. We need insights from both the top down and the bottom up. I would be exceedingly pleased if undergraduate and graduate students found the book to be a useful compass, pointing, at least in a general sense, in the direction that new research in evolutionary ecology might most fruitfully proceed.
Acknowledgments
Much of this book was written while I was on a sabbatical visit to Monash University in Melbourne, Australia, for the first part of 2018, hosted by my friend and colleague Damian Dowling. The Dowling lab has emerged as one of the most innovative research teams in the world, pushing forward the boundaries of understanding of mitonuclear ecology. Damian and his lab group could not have been more welcoming and encouraging as I struggled to write some of the most challenging chapters of the book. Damian and the Dowling lab read every chapter in early draft stages and
provided hugely valuable input. While in Australia, I also had extensive discussions and received critical feedback on my book from other colleagues, particularly Paul Sunnucks, Leo Joseph, and Chris Greening. Chris Greening is a brilliant biochemist with interests that range from the physical interaction of atoms to how climate might shape the ranges of Australian birds, and he did his best to coach me through the complexities of oxidative phosphorylation. Leo Joseph, curator of birds at the Commonwealth Scientific and Industrial Research Organisation (CSIRO) and an international expert on species boundaries in birds, critiqued the chapter on speciation. Paul Sunnucks showed me the potential impact that mitonuclear thinking can have on studies of phylogeography in the field as we trapped eastern yellow robins on his study sites. The faculty and students at Monash served as vital sounding boards for my ideas, and I am very grateful for the time and effort they invested in making my book better. In Australia I also went on a lecture tour, presenting my ideas at universities throughout Victoria, New South Wales, and the Capital District. The opportunity to vet my ideas in front of hundreds of biologists from diverse backgrounds helped me gauge how the ideas in my book were likely to be received. Thanks to all of my Australian colleagues who vetted my ideas. And finally, my most important first-draft readers were the students and postdocs in my own lab and in the lab of my wife and close colleague, Wendy Hood, at Auburn University. The Hill–Hood lab groups were key in helping organize and construct chapters, and Wendy was the great motivator for getting the project done. Other colleagues including Daniel Sloan, Justin Havird, Ron Burton, Ryan Weaver, Zhiyuan Ge, Nick Justyn, Matt Powers, Yufeng Zhang, Halie Taylor, Noel Park, Kyle Heine, Chloe Josefson, Kristjan Niitepold, Tori Andreasen, Rebecca Koch, Andrea Pozzi, Rebecca Vaught, Winston Yee, Vanessa Higham, Sean Layh, Magdalena Nystrand, Ilaria Venturelli, Ekta, Yoshana Fonseka, Venkatesh Radha, and Tara-Lyn Carter provided key feedback on various chapters. Sasha Pavlova and Hernan Morales provided unpublished data and figures.
The idea for writing this book emerged initially from a weekly reading/discussion group centered on Nick Lane’s book Power, Sex, Suicide: Mitochondria and the Meaning of Life that I organized in the Department of Biological Sciences at Auburn University in the fall of 2012. That discussion group was formalized into a graduate class, “Mitonuclear Ecology,” that I taught at Auburn in 2014 and 2016. Notes from my Mitonuclear Ecology class formed the foundation of this book. The students, postdocs, and faculty who participated in those classes played key roles in shaping my thinking about the coevolution and coadaptation of mt and N genes, and to the participants in those discussion groups I owe the impetus for creating this book.
1
The genomic architecture of eukaryotes
Life is a rejection of entropy. It succeeds by harnessing and focusing energy toward the maintenance of order and stability. The greater the complexity of the organism, the more energy that is required to hold back the surging tide of chaos. Hence, the story of life on Earth is fundamentally a story of location, extraction, and production of energy. The origin of complex life was enabled by a radical redesign of simple prokaryotic cells that created unprecedented opportunities for production of energy. The linchpin of this evolutionary innovation, which changed the nature of life on Earth, was the mitochondrion.
The ascension to complexity was not an inevitable endpoint in the evolution of life. Eukaryotic life emerged from exclusively prokaryotic world in which tiny life forms extracted energy from the environment with fantastic efficiency but gained little in size or overall complexity through 2 billion years of evolution (DeLong et al., 2010). With modest capacities for energy extraction, Lane (2015a) speculated that this exclusively prokaryotic world of small and relatively simple organisms may have persisted indefinitely. There seemed to be no escape from the perpetual race to replicate. But the improbable evolution of a new type of organism, which carried two genomes, provided the radical restructuring of cellular design that jarred life out of its perpetual race for more efficient energy extraction and faster replication speed. The delegation of life processes to two genomes was an inescapable necessity in the evolution of complex life, and it is in the improbable fusion of prokaryotic life to create eukaryotic life that our story begins.
Eukaryotic evolution
Eukaryotes are chimeras
There is now compelling evidence that the origin of eukaryotes, the origin of mitochondria, and the origin of complex life were one and the same event (Lane, 2015b; Martin et al., 2015). About 2 billion years ago, two prokaryotic lineages established an intimate symbiotic relationship, evolving over an unknown but presumably long period to become ever more interdependent. One of these partners was a prokaryote belonging to the life domain archaea and the other was a prokaryote belonging to the life domain bacteria, and in a pivotal event in the history of life on Earth, these two
prokaryotic cells fused to form an entirely new organism that would evolve into a novel life domain called eukaryota, the eukaryotes (Williams et al., 2013; Poole and Gribaldo, 2014). I use the term “fused” with purpose because this was not a simple exchange of genes. Rather, this was a joining of what had been two independent organisms into a wholly new type of organism in which both founders retained some degree of independent identity. This unlikely fusion was the starting point for the evolution of complex life on Earth, but it was far from a finished process at the moment when the archaeon and bacterium joined. The 2 billion years since this chimeric fusion has been one long negotiation between the united partners regarding how to partition cellular duties, who is responsible for what genes, and, above all, how to coordinate genomic products to enable system function. Like friends sharing an apartment, the relationship requires a foundation of cooperation, but conflict is never far away.
Martin and Müller (1998) proposed that the likely circumstance that gave rise to the archaeon/bacterial chimera was an intimate symbiosis between the two cell types. This theory, which has become known as the hydrogen hypothesis, proposes that the host cell (the cell that subsumed its partner cell) was an archaeon that derived energy by ingesting hydrogen and carbon dioxide and using these materials to produce methane and energy. This methane producer, or methanogen, required access to free hydrogen, which is scarce in most Earth environments. Fortuitously, hydrogen was the waste product of a second cell type, a bacterium that derived at least some of its energy from anaerobic metabolism of organic material to produce hydrogen and carbon dioxide. The waste of one partner was the energy source for the other and so they were naturally drawn together into a tight symbiosis. Eventually, the bacterium literally moved inside the archaeon and became an internal organelle.
What followed was a series of cataclysmic events within the new chimeric cell as the components of two very different prokaryotic organisms reorganized into a single life entity. The fascinating details of this restructuring of the proto-eukaryotic cell into a true eukaryotic cell are told in brilliant prose in Nick Lane’s books Power, Sex, Suicide: Mitochondria and the Meaning of Life and The Vital Question: Energy, Evolution, and the Origins of Complex Life (Lane, 2005, 2015a), as well as in dozens of articles in technical journals where data and ideas are presented one piece at a time. For the purposes of this book, the essential information is that the bacterium became the mitochondrion of eukaryotic cells and the archaeon became the “host cell” such that its genetic material evolved into the nuclear (N) genome (Sagan, 1967; Gray, 2012). There is strong evidence that this archaeon/bacterium chimeric fusion occurred only once and that all eukaryotes—animals, plants, fungi, protozoa, slime mold, brown algae—share a common ancestor (Williams, 2014; Derelle et al., 2015). In stating that all eukaryotes evolved from a common ancestor and hence that the chimeric fusion of an archaeon and a bacterium occurred only once, I am in no way suggesting that life succeeded in its initial run at this unlikely fusion and restructuring of cells. Given all of the challenges associated with this new architecture, which I review in the first five chapters of this book, one can imagine that the experiment started and failed an incalculable number of times before a thousand fortuitous events fell in sequence and
the union succeeded. Keep in mind there is more than a billion years between the rise of bacteria and archaea and the appearance of the first eukaryote, and a billion years provides ample time for extremely improbable events to occur.
From the initial fusion of archaeon and bacterial cells to form a proto-eukaryote, the archaeon genome existed as one (or two in diploids) copy per cell and the bacterial genome existed as multiple copies. In discussing the origin of eukaryotes from the initial bacterium/archaeon fusion, I will focus on the genomes of the two organisms and in so doing I will set the stage for the central theme of the book: coadaptation of mitochondrial (mt) and N genomes. Because mt genes code exclusively for protein subunits of the electron transport system (ETS) or for components of the transcriptional, translational, and replicative machinery needed to produce ETS subunits, it is essential to start with a brief review of how eukaryotic cells produce ATP via aerobic respiration.
OXPHOS and the electron transport system
Most eukaryotes acquire the energy needed for life processes at least partly through aerobic respiration. The first two phases of cellular respiration, glycolysis and the citric acid cycle, occur in the cytoplasm of the cell and in the mitochondrial matrix, respectively (Figure 1.1). These initiating steps in the process of cellular respiration take food
Figure 1.1 A simplified overview of key features of mitochondrial and cellular respiration. The inner mitochondrial membrane divides the cell into the intermembrane space, which lies between the inner and outer mitochondrial membranes, and the mitochondrial matrix, which is the chamber inside the inner mitochondrial membrane. Mitochondrial DNA exists within the mitochondrial matrix. The complexes of the electron transport system are embedded in the inner mitochondrial membrane, forming a critical conduit between the intermembrane space and the mitochondrial matrix. Glycolysis, the first stage of cellular respiration, occurs outside of mitochondria in the cytosol. The second stage of aerobic respiration, the citric acid cycle, occurs in the mitochondrial matrix. OXPHOS is enabled by the electron transport system.
Outer mitochondrial membrane Intermembrane space
Mitochondrial matrix
Electron Transport System (ETS) Complexes
molecules like glucose and break them down to release a small amount of ATP and a substantial amount of the energy-rich coenzymes NADH or FADH2. Despite the appearance that most of the molecular decomposition of food molecules is finished by the end of the citric acid cycle when NADH and FADH2 are produced, about 90 percent of the energy available in a glucose molecule is still retained in NADH or FADH2. This huge remaining energy pool is captured via the process of oxidative phosphorylation (OXPHOS), which is enabled by a series of integrated protein complexes called the electron transport system (ETS) (Box 1.1). The first four complexes of the ETS are
Box 1.1 Ox Phos Wars
By the middle of the twentieth century, cell biologists had deduced the basic chemistry of cellular respiration and identified ATP as the key molecule used by organisms to store energy in a usable form. Biochemistry, like all chemistry, is founded on stoichiometry— balancing the precursors and products in reactions. In basic chemistry, atoms are never created or destroyed; stoichiometry works strictly with integers. When a scientist balances a chemistry equation, he or she is precluded from ending with, say, 73 percent of a hydrogen atom. Atoms are recombined as whole units. So, it was perplexing to say the least that the stoichiometry of cellular respiration never produced integers—it invariably produced fractions of integers. The persistent failure of the reactions driven by the ETS to stoichiometrically balance in experiments indicated that something very fundamental was missing from the theory of cellular respiration.
The paradox of equations that would not balance lasted for more than 20 years until Peter Mitchell solved the problem by hypothesizing that production of ATP involved the creation of an electrochemical gradient across a membrane (Mitchell, 1961). This process of converting food energy into a proton-motive force and then using a membrane potential as the energy source for ATP production lifted the restriction of stoichiometric precision. Protons could be released from the gradient at variable rates enabling ATP to be created in nonstoichiometric ratios.
Like many ideas that represent huge and non-intuitive advances in fields of study, Mitchell’s chemiosmosis theory was not embraced quickly or enthusiastically by most of his colleagues. As a matter of fact, arguments about the mechanism by which oxidative phosphorylation generated ATP were so vitriolic that the debate became known as the Ox Phos Wars (Prebble, 2002). Mitchell was subjected to substantial ridicule in the years following the publication of his brilliant hypothesis, and it was not widely accepted as correct for years after it was initially published. In the end, Mitchell received the Nobel Prize in chemistry for his discovery, so when recognition came it came with the highest honor in science.
Studies of mitonuclear ecology build from the insights of Peter Mitchell. The chemiosmosis hypothesis of cellular respiration helped refine understanding of the ETS—the large protein complexes that use energy from electrons to pump protons across a membrane and to create a chemiosmotic energy potential (Slater, 2003). It created the foundation on which the significance of mitonuclear coadaptation and coevolution could be understood. The discovery of chemiosmosis as the basis for energy production in living organisms required a tremendous intellectual leap and great courage by Mitchell, and it stands as one of the most significant discoveries in the history of science.
often referred to as the electron transport chain or respiratory chain, and I will use these terms periodically in this book to refer to complexes that transfer electrons.
The ETS is a set of large and complex proteins that receive high-energy electrons from either NADH or FADH2 and use the energy of these electrons to pump protons across the inner mitochondrial membrane, from the mitochondrial matrix (mitochondrial core) to the intermembrane space. Pumping protons across the membrane creates a net positive charge in the intermembrane space relative to the matrix (Figure 1.2). The membrane potential is the difference in electrical charge created by an excess of protons on one side of the inner mitochondrial membrane. Maintaining this membrane potential is critical for cell survival. In a healthy, properly functioning cell, the gain of protons from active pumping is counterbalanced by discharge of protons back across the membrane. Discharge of protons is typically through ATP synthase (Complex V), which phosphorylates ADP to produce ATP from the energy recaptured from protons as they move back across the membrane (Figure 1.2). Thus, food energy is used to “crank” ATP synthase (the molecule actually pivots on a molecular axis) to generate ATP. It is hard not to use the analogy of the energy potential of water held behind a dam that turns a wheel as it flows to a point below the dam, but the physical mechanisms involved with the flow of water driven by gravity and the flow of protons across an electrochemical gradient are distinct.
In its textbook description, the ETS is composed of five protein complexes designated as: Complex I, II, III, IV, and V (Figure 1.2). More technically, Complex I is
Figure 1.2 A stylized illustration of key elements of the electron transport system. Depicted is a cross section of a mitochondrion indicating the relative position of components. The top right cluster with Complexes I, III, and IV illustrates the primary path of electrons from NADH to Complex I. The bottom left cluster with Complexes II, III, and IV illustrates an alternative route of electrons from FADH2 to Complex II. Both routes end with Complex V converting the proton motive force to ATP.
NADH dehydrogenase; Complex II is succinate dehydrogenase; Complex III is coenzyme Q–cytochrome c reductase; Complex IV is cytochrome c oxidase; and Complex V is ATP synthase. Complex V is sometimes not included as part of the ETS because it does not transport electrons. Because ATP synthase is the terminal enzyme in the transfer of energy from electrons to ATP, I include ATP synthase when I refer to the ETS. In addition to the large protein complexes that span the inner mt membrane, there is a single-unit protein, cytochrome c, that transports electrons from Complex III to IV and a vitamin-like quinone, coenzyme Q, that transports electrons between Complexes I or II and Complex III.
The I to IV numbering of the components of the ETS suggests a linear series of actors, but this is not the case. There is core electron input from NADH in Complex I, which is the starting point for most energy production. In addition, there are side electron inputs from three flavoprotein complexes, namely Complex II, electrontransferring flavoprotein dehydrogenase, and glycerol 3-phosphate dehydrogenase (Frerman and Goodman, 2001; Ishizaki et al., 2005). The electrons from these two unequal sources converge in Complex III for most eukaryotes (Figure 1.2). But even the necessity of Complexes III and IV cannot be stated as a universal generality because in plants and some other eukaryotes, an alternative oxidase (AOX) enables the reduction of oxygen without Complex III or IV (Saha et al., 2016). Thus, for most eukaryotes and in all of the discussion in this book, there are four core complexes of the ETS system—Complex I, III, IV, and V—plus three side inputs, including Complex II.
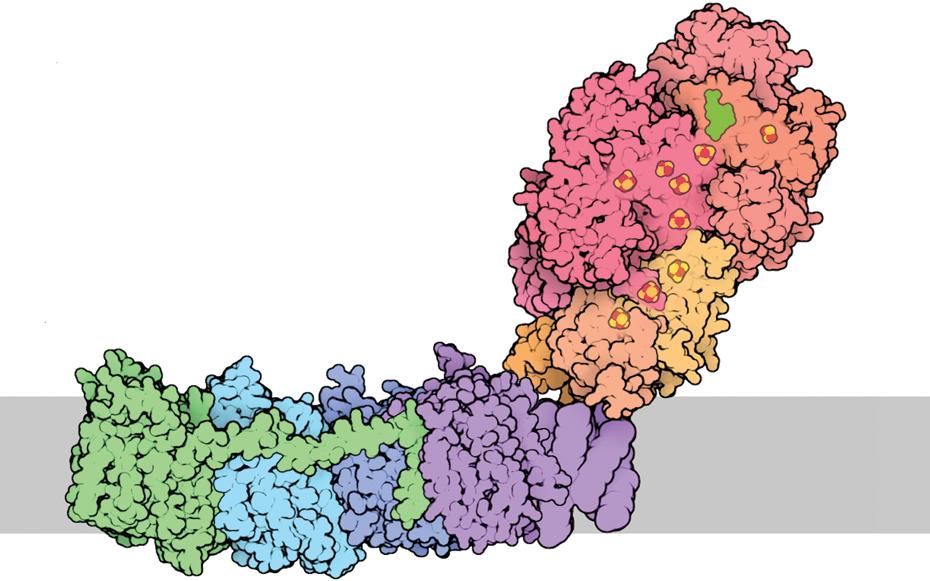
The large components of the ETS are called “complexes” because they are composed of multiple, independently coded protein subunits. For instance, the mammalian Complex I is assembled from about forty-six protein subunits (Hirst, 2013). Each of these protein subunits is a three-dimensional structure that is complicated both in shape and in the distribution of reactive sites and charges. To create a functional ETS complex, each subunit must have a precise three-dimensional configuration that enables it to join and interact correctly with its neighboring subunits (Figure 1.3). The distribution of reactive sites and electrical charge across the structure of any given subunit must be complementary to all neighboring subunits. There must also be complementarity between cytochrome c and both Complex III and Complex IV for efficient transfer of electrons. Mutations that lead to changes in a component of an ETS complex or cytochrome c can compromise the functionality of a complex and in turn interfere with the functioning of the entire ETS. Such subunit incompatibility is the topic of Chapter 2 and will be the focus of much of this book.
Massive genomic restructuring
Nature favors economy over excess (Wolf and Koonin, 2013). When two organisms exist in a tight symbiosis, it is common for genes whose function is duplicated between the two partners to be lost in one of the symbionts (Moran, 2003, Moran et al., 2008; McCutcheon and Moran, 2012). For instance, the bacterium Carsonella
Figure 1.3 A three-dimensional reconstruction of a bacterial Complex I molecule with fourteen protein subunits, each shaded a different color. The position of the membrane in which the molecule is embedded is shown in gray. This model is presented to emphasize the intricate three-dimensional fit among subunits that is necessary to produce a functional complex. The human Complex I molecule has forty-six subunits—more than three times the structural complexity of the illustrated bacterial complex. Image from the Research Collaboratory for Structural Bioinformatics Protein Data Bank (RCSB PDB) “Molecule of the Month”: Inspiring a Molecular View of Biology (Goodsell et al. 2015).
ruddii is an ancient symbiont of the sap-feeding insect Pachypsylla venusta and it has lost so many genes through eons of coevolution with its host that it is arguably now an organelle (Sloan et al., 2014a). Carsonella ruddii has lost not only genes whose functions are duplicated by host genes, but has also transferred some genes that are essential for vital biosynthetic pathways (Sloan et al., 2014a). The dynamics of genome reduction in obligate symbionts provide insights into the evolution of the N and mt genomes in early eukaryote evolution (Moran and Wernegreen, 2000; Andersson et al., 2003; Rand et al., 2004).
Once the primordial eukaryote emerged from the fusion of two formerly independent cells, genes that existed as two versions that duplicated the same function in the two entities were reduced to one version in the unified organism (Timmis et al., 2004). In most cases, the version of the gene that was retained was located in the N genome because, from the beginning, genes in the nucleus existed as one copy (technically two copies in diploids) per cell, while genes in the mt genome existed as many copies in the multiple mitochondria inside the cell. It was fundamentally more efficient to
maintain most genes in the unique central genome of the cell rather than as multiple copies in mt genomes, and the nucleus quickly became the information center of the eukaryotic cell (Adams and Palmer, 2003). Moreover, as the eukaryotic genomes evolved, the N genome became sexual and evolved the capacity for recombination (see Chapter 5 for justification of the assumption that mitochondria evolved before sexual reproduction). The N genome also typically had lower mutation rates compared with the mt genome to which it was matched (Lynch, 1997) and, unlike the mt genome, the N genome avoided Hill–Robertson effects via recombination (discussed in detail in Chapter 5). Thus, the nucleus became the better depository for genomic information (Allen and Raven, 1996; Blanchard and Lynch, 2000). The gene copy that was retained in the nucleus was frequently of archaeon origin, because such genes were already located in the N genome, but sometimes the bacterial version of the gene was moved to the nucleus and the archaeon version was lost (Gray, 2012). Interestingly, even though there is now broad consensus that the mitochondrion evolved from a single bacterial ancestor, only about 14–16 percent of the proteins that make up the mitochondrion of a modern eukaryote seem to be of bacterial origin (Gabaldón and Huynen, 2004). Components of the bacterial partner have been modified, added to, and replaced in the course of evolution from free-living cell to mitochondrion (Gray, 2015).
The culling of duplicate genes from the mt genome resulted in a substantial reduction of mt genome size (Gray, 2012) (Figure 1.4), but shrinking of the mt genome was not limited to the removal of duplicate genes. Many unique mt genes that were critical for mitochondria-specific function were also transferred to the nucleus (Adams and Palmer, 2003). This transfer of functional mt genes to the nucleus necessitated that, constantly through the life of every eukaryotic organism, numerous proteins synthesized outside the mitochondrion had to be transported into the mitochondrion (Bolender et al., 2008; Calvo and Mootha, 2010). Very rapidly in the evolution of eukaryotes, nearly all of the genes that had originally comprised the bacterial genome were deleted or moved to the nucleus (Figure 1.4). Most modern species of alpha-proteobacteria, which is proposed to be the class of bacteria from which mitochondria evolved (Lang et al., 1999), have genomes with about 2000–4000 genes (Koonin and Wolf, 2008); in contrast, the mt genomes of bilaterian animals have thirty-seven genes (see Box 1.2 for a discussion of variation in mt genome size). Thus, about 99 percent of the genes that were present in the bacterial partner that evolved into the mitochondrion in the early evolution of eukaryotes were lost or moved to the nucleus (Figure 1.4). This process of gene transfer was so nearly complete that an obvious question is: why weren’t all genes moved out of the mitochondria (Kleine et al., 2009)? Why retain that last vestige of mt genes? This entire book is about the enormous consequences of the retention of those few dozen mt genes for the evolution of complex life, so it is critical to consider why such gene retention was not a failure of an evolutionary purge, but rather an absolutely necessary feature of the genomic structure of complex life forms that derive energy from aerobic respiration.
Ancestral Archaeon: ~3000 genes
Before chimeric fusion
Archaeon Bacterium
Genomic rearrangements
Evolution of many novel genes
Gene transfer
Gene loss
Ancestral Bacterium: ~3000 genes
Nucleus ~20,000 genes
Eukaryote
Mitochondrion: 37 genes
Figure 1.4 A general summary of key events in the genetic restructuring in the early evolution of eukaryotes. The three panels cover an evolutionary period of unknown duration beginning about 2 billion years ago. In the evolution of eukaryotes, many genes in the original bacterial genome were transferred to the nucleus or deleted. The number of genes in the former archaeon genome increased by more than an order of magnitude.
The mitochondrial genome
The retention of a mt genome
In the formulation of early hypotheses for why a small number of genes were retained in mitochondria, a basic assumption was that an optimal endpoint was zero genes in the mitochondrion (Björkholm et al., 2017). Researchers therefore proposed that loss of mt genes was an ongoing process that hadn’t had time to reach completion, and they sought to understand the constraints on the process of gene transfer. Recent papers considering evolutionary constraints on the complete loss of mt genes have
focused on difficulties related to protein transport. Because mt-encoded proteins are highly hydrophobic, they may be difficult to move across mt membranes and hence difficult to get into mitochondria if they are encoded by N genes (Claros et al., 1995; Björkholm et al., 2015). This mechanistic constraint is proposed as the reason that genes were retained in the mitochondrion. However, there are examples of hydrophobic proteins being moved into organelles and particularly chloroplasts (Allen, 2003). Other hypotheses for how constraints can explain retention of a mt genome have also been proposed (summarized in Allen, 2003).
Even though hypotheses focused on the constraints of moving hydrophobic proteins from the mt genome to the N genome remain viable explanations for the current genomic architecture of eukaryotes, I will proceed from the argument that retention of mt genes is not the result of a failed purge, but rather that in the evolution of eukaryotes there was selection to retain genes in the mitochondria. This view is supported by the observation that all eukaryotes that produce energy via aerobic respiration retain a common core set of mt genes (Boore, 1999; Björkholm et al., 2015) (Box 1.2). In addition, the loss of mt genes was completed in eukaryotic lineages that lost the need for a functional ETS (Smith and Keeling, 2015). Thus, in all organisms that retained aerobic respiration, core components of the mt genome were retained; when aerobic respiration was lost, so were genes remaining in mitochondria.
Co-location for redox regulation (CORR)
A key assumption in this book is that the mitochondria of most eukaryotes retained genes because genes in the mitochondria provide critical functions that enable complex life. The key to this assumption is the current leading hypothesis to explain the retention of a small genome in mitochondria: the co-location for redox regulation (CORR) hypothesis (Allen, 1993, 2003, 2017). This hypothesis states explicitly that mt genes are critical to complex life forms that derive energy from OXPHOS. The central tenet of the CORR hypothesis is that individual mitochondria must retain transcriptional and translational control of core ETS proteins so that individual mitochondria can rapidly respond to the current conditions of their inner mitochondrial membranes (Sirey and Ponting, 2016). This hypothesis focuses on the massive levels of energy needed to power complex life, with many trillions of mitochondria in billions of individual cells each creating an electrical potential across inner mitochondrial membranes that is equivalent to the energy in a bolt of lightning (Lane, 2006; Lane and Martin, 2010). The CORR hypothesis proposes that retention of mt genes was necessary to maintain control of such a massive energy generation system.
Efficient OXPHOS via a highly functional ETS is the key to eukaryotic fitness. For the ETS to work efficiently in enabling OXPHOS and capturing energy from the membrane potential, there must be precisely the correct numbers and ratios of ETS complexes to meet the demands of energy production (Lane, 2005). If the ratio of complexes is incorrect or if there are insufficient total ETS complexes, then electrons at one complex in the chain are frequently left with no empty positions in the next complex in the chain as they move toward Complex IV where the electrons are finally passed to oxygen to produce water. These stalled electrons dissipate their energy by
Box 1.2 Variation in mitochondrial gene content
Mitochondrial DNA codes for three general classes of genes: ribosomal RNA, transfer RNA, and protein-coding. Although there is variation among major clades of eukaryotes, and especially among single-celled eukaryotes, the gene content of mt genomes is remarkably conserved among all aerobic eukaryotes. Mitochondrial genomes always include genes that code for proteins that carry out core function of ETS complexes but never enough genes for complete ETS function. In all aerobic eukaryotes, mt DNA codes for at least some part of the mechanism for transcription and translation of ETS proteins. Even if none of the proteins needed for transcription is mt-encoded, as is true for many eukaryotes, the nucleotide sequence of regulatory sites in the mt DNA must be compatible with N-encoded transcriptional machinery (Ellison and Burton, 2010). The universal contribution to translation by the mt DNA of all aerobic eukaryotes is the small and large ribosomal RNAs (rRNAs). Some, but not all, eukaryotes also code for ribosomal proteins. Most eukaryotes also code for transfer RNAs (tRNAs), but these are lost completely in a few protozoa, which rely on tRNAs from the nucleus (Burger et al., 2003; Castellana et al., 2011) (Figure Box 1.2).
Among bilaterian animals, which will be the focus of a disproportionately large part of this book, there is little variation in the gene content of mt DNA. There are thirty-seven genes that include two ribosomal RNAs, a full complement of twenty-two tRNAs, and thirteen proteins that provide core subunits of Complex I, III, IV, and V of the ETS (Castellana et al., 2011).
OXPHOS proteins
Ribosomal RNAs
Transfer RNAs
Ribosomal proteins
Protein import & maturation
RNA maturation
Transportation
Figure Box 1.2 A graphical representation of the gene content of mitochondrial DNA from diverse eukaryotic lineages. The red box bounds the core mitochondrial gene products common to all eukaryotes that derive energy via OXPHOS. Figure adapted from Burger et al. (2012).
side reactions with oxygen, resulting in the double whammy of less ATP generated and more cellular damage from free radicals (Lane, 2011c). If things go very wrong with the inner mitochondrial membrane potential, then cell death is triggered and the entire operation is shut down and disassembled (Ly et al., 2003). Only when the number and ratio of ETS complexes matches the current need of the system do electrons flow at an optimal rate producing energy efficiently and releasing few free radicals in side reactions (Allen, 2003; Lane, 2014). The CORR hypothesis proposes that the OXPHOS via the ETS can only be properly controlled if each mitochondrion has the capacity to regulate production of ETS subunits, and such control is only possible if each mitochondrion has its own set of genes for ETS components.
The challenge for eukaryotic organisms is that the conditions within one mitochondrion in a cell are not necessarily the same as conditions within other mitochondria within the same cell. There are too many small and ever-changing variables that affect the immediate conditions within a given mitochondrion. This means that it is not possible to properly regulate the ETS and maintain efficient OXHPOS within numerous individual mitochondria by regulating gene expression entirely within the N genome (Lane, 2005, 2015a; Lane and Martin, 2010). Attempting to control respiration in hundreds or thousands of mitochondria within a cell from a central command center would be like trying to control the boilers in hundreds of schools within a large school district from a central administrative office. Each boiler has to maintain an appropriate steam pressure for efficient function but each boiler is subject to untold variables as thermostats are changed, windows are opened or closed, outside temperatures change, and so forth. If boilers were checked once per day by administrators in the central office, individual systems would be perpetually maladjusted. On the other hand, if each boiler room was staffed by a technician who watched the dials and adjusted the system according to immediate conditions in that particular school, then each individual system would run much more efficiently and the district as a whole would function well.
According to the CORR hypothesis the mitochondria in all eukaryotes that engage in OXPHOS retained a small set of genes to enable within-mitochondrion transcriptional or translational regulation of proteins in the ETS complexes in response to the immediate local conditions (Allen, 2003). Without such rapid local responsiveness, membrane potential within an individual mitochondrion can drop, leading to cessation of ATP production, release of free radicals and other molecules that ultimately signal cell death. The CORR hypothesis proposes that all aerobic eukaryotes retained a core set of mt genes as a fundamental necessity of mitochondria-based respiration (Lane and Martin, 2010).
An observant reader might note that only a minority of ETS subunits are encoded by mt genes. For instance, there are approximately ninety-four subunits making up the five complexes of bilaterian animal ETS and only thirteen of these subunits are contributed by the mt genome. Thus, the great majority of proteins that create the ETS are encoded in the nucleus. Given this arrangement, how can transcriptional or translational regulation of thirteen out of ninety-four subunits enable mitochondria to control overall production of ETS complexes? Many details of the mechanisms of complex assembly remain to be worked out (Guaras and Enriquez, 2017), but it is almost surely no coincidence that the subunits that are coded for by mt DNA form
the proton-translocating center of the complexes that localize within the inner mt membrane (Allen, 2015). Lane (2005, 2014) hypothesized that N-encoded respiratory subunits assemble around core mitochondria-encoded subunits once they are targeted to the membrane, and this hypothesis is gaining empirical support (Formosa et al., 2017). The production of N-encoded ETS subunits is dictated by the redox state of the entire cell—the average state of all mitochondria in the cell. But the specific rate of assembly of ETS complexes is dictated by mt genes that are responsive to the immediate conditions within each mitochondrion within the cell. According to this idea, N genes follow the lead of mt genes in assembling complexes. Control of mt genes therefore does provide control of complex assembly.
This proposed scenario for the assembly of ETS complexes makes the point for why it is so difficult for evolution to purge the genes for key membrane-anchored ETS subunits from the mt genome. These protein products are the enablers of locale transcriptional and translational regulation. Moreover, the signaling network that guides responsiveness to the state of the mitochondrion acts through transcription, translation, and replication of mt genes. Thus, it was essential to retain in the mt genome the genes for membrane-embedded ETS proteins as well as key components of the transcription, translation, and replication systems.
To summarize the logic for why two genomes were so important to the evolution of complex life: complex life forms require large genomes that code for huge quantities of diverse proteins. Sufficient energy to power such complexity is made possible by a cell design with a single large centralized genome and many independent mitochondria that act as power stations to support the enormous energy demands of the production of proteins encoded by the N genome (Lane and Martin, 2010). Such a cellular design absolutely necessitated the retention of a mt genome for autonomous control of the ETS by each mitochondrion to maintain the stability of the respiratory process (Lane and Martin, 2010; Lane, 2011c). The train of deductions proceeds as follows: complexity requires a large genome; a large genome requires a lot of energy; massive energy production is only possible via the combined output of many mitochondria; and control of such a power system necessitates independent genomes in each mitochondrion. As I will argue throughout this book, the necessity of coadaptation between the N and mt genomes led to the evolution of sex, two sexes, speciation, sexual selection, and adaptation.
The endpoint of gene transfer
As a result of the retention of core components of the ETS and the transcriptional and translational mechanisms needed to produce mt-encoded proteins, the phenotype of mitochondria is a product of two genomes (Figure 1.5). All of the gene products of the mt genomes—thirty-seven genes in most bilaterian animals and as many as sixty-six genes in some jakobid protozoa—stay at home and create mitochondrial form and function. Mitochondria are hugely complex organelles that complete a long list of functions that are essential for eukaryotic life (Prasai, 2017). A few dozen genes is not nearly enough to code for a functional mitochondrion; only about 2 percent of the proteins that make up a mitochondrion are encoded in the mt genome (Gray, 2015).