Oxford University Press is a department of the University of Oxford. It furthers the University’s objective of excellence in research, scholarship, and education by publishing worldwide. Oxford is a registered trade mark of Oxford University Press in the UK and certain other countries.
Published in the United States of America by Oxford University Press 198 Madison Avenue, New York, NY 10016, United States of America.
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, without the prior permission in writing of Oxford University Press, or as expressly permitted by law, by license, or under terms agreed with the appropriate reproduction rights organization. Inquiries concerning reproduction outside the scope of the above should be sent to the Rights Department, Oxford University Press, at the address above.
You must not circulate this work in any other form and you must impose this same condition on any acquirer.
CIP data is on file at the Library of Congress
ISBN 978–0–19–067641– 4
9 8 7 6 5 4 3 2 1
Printed by Sheridan Books, Inc., United States of America
I dedicate this book to my lovely wife Jamie, and I express my heartfelt appreciation for her love, encouragement and support for this work and throughout my career.
Harvey B. Lillywhite
I dedicate this book to my lovely wife Eliana for having been a great source of inspiration and support throughout this work
Marcio Martins
Foreword
Acknowledgments
List of Contributors xv
1. Ecology of Snakes on Islands—Marcio Martins and Harvey B. Lillywhite 1
2. Isolation, Dispersal, and Changing Sea Levels: How Sea Kraits Spread to Far-Flung Islands—Harold Heatwole 45
3. Terrestrial Habitats Influence the Spatial Distribution and Abundance of Amphibious Sea Kraits: Implication for Conservation—Xavier Bonnet and Fran çois Brischoux 72
4. Physiological Ecology of Sea Kraits Inhabiting Orchid Island, Taiwan—Ming- Chung Tu and Harvey B. Lillywhite 96
5. The Queimada Grande Island and Its Biological Treasure: The Golden Lancehead—Marcio Martins , Ricardo J. Sawaya , Selma Almeida- Santos , and Otavio A. V. Marques 117
6. Pleasure and Pain: Insular Tiger Snakes and Seabirds in Australia— Fabien Aubret 138
7. The Eyes Have It: Watching Treeboas on the Grenada Bank— Robert W. Henderson
8. The Ecology and Conservation of the Milos Viper, Macrovipera schweizeri G öran Nilson 181
9. The Unique Insular Population of Cottonmouth Snakes at Seahorse Key—Harvey B. Lillywhite and Coleman M. Sheehy , III
10. Living Without a Rattle: The Biology and Conservation of the Rattlesnake, Crotalus catalinensis, from Santa Catalina Island, Mexico—Gustavo Arnaud and Marcio Martins
11. Decline and Recovery of the Lake Erie Watersnake: A Story of Success in Conservation—Richard B. King and Kristin M. Stanford
12. Defending Resources on Isolated Islands: Snakes Compete for Hatchling Sea Turtles—Akira Mori , Hidetoshi Ota , and Koichi Hirate
13. Islands in the Sky: Snakes on South American Tepuis—D. Bruce Means and C ésar Barrio- Amor ós
FOREWORD
“Islands” and “snakes” are two words that evoke a powerful sense of discovery and adventure. Islands are realms of endemism and novelty, and their exploration has informed some of the most fundamental ideas in the history of biology. Snakes reflect the mystery and beauty in nature, and our hardwired fascination with them reminds us of our intimate connection with it. Editors Harvey Lillywhite and Marcio Martins reveal this world by drawing on their diverse collaborations and collective decades of scholarship and passion.
Nineteen accomplished snake biologists, alongside Lillywhite and Martins, have contributed chapters that together cover the ecology, behavior, evolution, and conservation of snakes on islands. Each chapter is illustrated with color photographs of spectacular snakes and their island habitats. These range from bird-eating tree boas in the Caribbean to amphibious sea kraits spanning Taiwan and New Caledonia and castaway Australian tiger snakes. The authors go far beyond the existing scientific literature by allowing the reader privileged insight into the passion and process behind their discoveries. The entertaining anecdotes shared in each chapter show that new questions and new knowledge are gained from lifelong curiosity and dedication, innovative thinking, and also some degree of risk-taking. Furthermore, by reflecting on their closest professional collaborations and enriching interactions with local communities, the authors reveal the human side of scholarship. Books such as this are vital for stimulating public enthusiasm for science and conservation.
Islands and Snakes: Isolation and Adaptive Evolution is timely. Island environments are under threat from development, rising sea levels, and an increasing incidence of invasive species. And biologists are racing against time to discover what species live on islands, the crucial roles they play there, and how they have adapted to island life. By relating ecological and evolutionary insights gained from field studies of snakes on islands, this book will no doubt inspire numerous new research and conservation initiatives. Islands and Snakes: Isolation and Adaptive Evolution is a must-have for students,
x • Foreword
biologists, geographers, and anyone who values fragile island environments and their unique biodiversity.
Kate Sanders
Adelaide, Australia
August 2018
PREFACE
During the fall of 2013, we taught a seminar course together on island ecology at the University of Florida, and the genesis of ideas for this book was in large measure an outgrowth of this class. At the beginning, we had intended to write a review paper that covered both historical and contemporary aspects of the biology of snakes on islands. Then, we scrapped this idea and replaced it with a goal of producing an academic book that focused on aspects of island ecology and biogeography as viewed through the lens of the many studies in which snakes have been a biological focus of such investigation.
Islands have been appreciated as natural “laboratories” for investigations of ecology, biogeography, and evolutionary biology since the time of Wallace, who dedicated a large amount of his writings to islands, including his seminal work Island Life, and Darwin, who was profoundly affected by his observations in the Galapagos Islands. In the 1960s, MacArthur and Wilson produced an important and influential theoretic framework for subsequent investigations of biodiversity and dynamics of insular biogeography. Since MacArthur and Wilson’s pioneering efforts, a robust literature on insular ecology and biogeography has continued to grow, and understanding the successful existence and adaptations to conditions on islands has advanced. Various investigators have extended earlier theoretic studies to increase understanding of important phenomena such as adaptive radiation, energetics, paleogeography, plasticity of colonizing biota, trophic changes, morphological evolution, and climate change.
For reasons of practicality and personal interests, various specific elements of biota have been investigated as model organisms for clarifying insights regarding particular features of island ecology. Reptiles on various islands have replaced endotherms as primary herbivores and top carnivores. Because of ectothermy, reptiles have advantages over endotherms in exploiting scarce resources in circumstances that are challenging to the success of birds and mammals. Snakes are known to be very successful colonizers of islands, and
roughly 60% of literature on insular squamate reptiles deals with snakes. Indeed, studies of snakes have contributed much to our understanding of insular ecology, and these vertebrates are important subjects for investigating questions that might be difficult to approach in other systems. Details concerning the reasons why snakes have been successful in living on islands may be found in Chapter 1.
We have been fortunate to observe snake populations on many islands throughout the world, including key locations in South America, the Gulf of California, Taiwan, Australia, and the tropical Pacific. In many instances, snakes on islands occur in amazing numbers and are often a dominant aspect of the local fauna. To produce this book, we have recruited authors from among authorities throughout the world who have focused influential studies of snakes that occupy interesting and important systems on various islands or archipelagos. As a concluding chapter, we have included studies of the spectacular tepuis that comprise an exceptional example of “ecological islands” in South America. All of the various authors provide entertaining narratives of the system they studied, woven as a fabric with solid empirical information, scientific theory, and personal insights regarding ecological and evolutionary principles as revealed by spectacular snakes and their adaptations to living on islands.
Harvey B. Lillywhite Marcio Martins
ACKNOWLEDGMENTS
We are grateful to the many persons who have made this book possible, especially our families, the various authors who have contributed thoughtful and stimulating chapters, numerous colleagues, reviewers, and others who have encouraged our adventures in science, including numerous visits to exciting islands. We also thank the editors and production staff at Oxford University Press for their professional guidance and assistance throughout this project. We hope that readers will find the enjoyment and satisfaction of reading this book that we have intended for them to discover.
CONTRIBUTORS
Selma Almeida- Santos
Laboratório de Ecologia e Evolução
Instituto Butantan
São Paulo, Brazil
Gustavo Arnaud
CIBNOR
La Paz, Mexico
Fabien Aubret
Station d’Ecologie Théorique et Expérimentale de Moulis
CNRS
Moulis, France
School of Molecular and Life Sciences
Curtin University, Australia
César Barrio-Amorós
Doc Frogs Expeditions
San Isidro del General, Costa Rica
Xavier Bonnet
CEBC- CNRS
Villiers en Bois, France
François Brischoux
CEBC- CNRS
Villiers en Bois, France
Harold Heatwole University of New England
Armidale, New South Wales, Australia
North Carolina State University Raleigh, North Carolina, USA
Robert W. Henderson
Section of Vertebrate Zoology
Milwaukee Public Museum Milwaukee, Wisconsin, USA
Koichi Hirate
Okinawa Prefectural Sea Farming Center Okinawa, Japan
Richard B. King
Department of Biological Sciences and Institute for the Study of the Environment, Sustainability, and Energy
Northern Illinois University DeKalb, Illinois, USA
Harvey B. Lillywhite
Department of Biology University of Florida
Gainesville, Florida, USA
Otavio A. V. Marques
Laboratório de Ecologia e Evolução
Instituto Butantan
São Paulo, Brazil
Marcio Martins
University of São Paulo
São Paulo, Brazil
D. Bruce Means
Coastal Plains Institute and Land Conservancy and Florida State University
Tallahassee, Florida, USA
Akira Mori
Department of Zoology
Graduate School of Science
Kyoto University Kyoto, Japan
Göran Nilson
Göteborg Natural History Museum
Göteborg, Sweden
Hidetoshi Ota
Institute of Natural and Environmental Sciences
University of Hyogo
Museum of Nature and Human Activities
Hyogo, Japan
Ricardo J. Sawaya
Centro de Ciências Naturais e Humanas
Universidade Federal do ABC
São Bernardo do Campo, Brazil
Coleman M. Sheehy, III
Division of Herpetology
Florida Museum of Natural History
University of Florida
Gainesville, Florida, USA
Kristin M. Stanford
F. T. Stone Laboratory
The Ohio State University
Put-in-Bay, Ohio, USA
Ming- Chung Tu
Department of Life Science
National Taiwan Normal University
Taipei, Taiwan
1
ECOLOGY OF SNAKES ON ISLANDS
Marcio Martins and Harvey B. Lillywhite
Introduction
Islands have been the subject of intense investigation—biologically and ecologically—since the time of Darwin and Wallace. Much research has focused on species assemblages and the dynamics of species richness on islands as well as other systems having geographic isolation and other characteristics similar to those of islands surrounded by water. An important theoretic model for insular biogeography was produced by MacArthur and Wilson (1963, 1967), and their work created a useful framework for subsequent investigations of biodiversity and its dynamics on islands (see reviews in Whittaker and Fernández-Palacios 2007; Warren et al. 2015; Santos et al. 2016; Patiño et al. 2017). Previously, insular faunas were regarded generally as either static or changing slowly and unpredictably due to environmental and climatic changes (Dexter 1978; Heaney 2000). However, the influence of more modern biogeographic theory enabled sometimes robust predictions of species richness in relation to the area of an island and its distance from a source of colonizing biota (MacArthur and Wilson 1967). Whether or not the biota of a given island or insular system is at “equilibrium” often remains debatable (Lomolino 2000; Warren et al 2015).
Since the 1960s, a robust literature on insular biology and ecology has continued to grow, with numerous investigators focusing on a variety of insular systems with attention to increasing detail concerning the requirements and dynamics of ecological factors that favor adaptation and successful existence on islands. Numerous questions are being addressed that cannot be explained by the existing theoretic models of insular biogeography (Gillespie
and Roderick 2002). Examples include (but are not limited to) questions regarding adaptive radiation (Losos 1998; Gavrilets and Losos 2009), paleogeography (Iturralde-Vinent and MacPhee 1999), energetics and energy resources (McNab 1994a, 1994b; Polis and Hurd 1996; Bonnet et al. 2002), climatic change (Bellard et al. 2013a, 2013b; Wetzel et al. 2013), life history characteristics of insular biota (Foufopoulos and Ives 1999), and anthropogenic influence (Steadman et al. 2005).
For many practical reasons (often with advantages), various investigators of the ecology of islands focus on taxonomic elements of fauna or flora with which they have familiarity or interest or that represent “model” systems due to a prior database of information or suites of characteristics appropriate to particular questions. Such approaches have intrinsic value and also provide important data for more inclusive investigations of biodiversity (Myers et al. 2000). Reptiles have been the frequent subject of investigations of vertebrate faunas on islands, where they are often “replacements” of endotherms as primary herbivores and top carnivores. The favorable circumstances for ectotherms versus endotherms on islands include low rates of energy expenditure relative to resources that are often scarce or limited on islands, dispersal abilities, and superior colonizing abilities. Rates of energy expenditure in terms of field metabolic rates of endothermic mammals and birds are approximately 12 and 20 times higher, respectively, than those of equivalent size, ectothermic reptiles (Nagy 2005). Reptiles, therefore, have a crucial advantage in exploiting scarce resources and building populations in circumstances that preclude, or severely challenge, the success of birds and mammals.
Snakes are very successful inhabitants of islands, and there is a very rich literature concerning the insular ecology of this group of vertebrates (Figure 1.1). An accounting of scientific articles using Google Scholar indicates that roughly 60% of the literature on islands and squamate reptiles (including Tuatara) deals with snakes. Thus, studies of snakes have contributed much to our understanding of insular ecology and are important subjects for attention to questions that might be difficult to investigate in other systems (discussed later). We emphasize that there are numerous reasons why snakes are important elements of insular biotas and play critical roles on numerous islands that can offer further insights for understanding ecology of islands. Some of the more important attributes of snakes related to successful “island living” are (1) ectothermy and comparatively low energy requirements; (2) attributes favoring abilities for overwater dispersal; (3) life history features favoring comparatively rapid population growth; (4) range of body sizes favorable for inhabiting even very small islands; (5) breadth and plasticity of diet, including
Figure 1.1 Examples of snakes having high significance with respect to insular ecology and biogeography. (a) Blue-banded Sea Krait (Laticauda laticaudata) from Orchid Island, Taiwan. Sea kraits are amphibious and spend variable amounts of time secluded on numerous small islands of the Indo-Pacific oceans (see Chapter 2). (b) Southeast Asian Bockadam (Cerberus schneideri) is an amphibious snake associated with estuarine habitats in Southeast Asia, including many areas of the Philippines. (c) Brown Tree Snake (Boiga irregularis) is widespread in the Oriental and Oceanian regions, occurring naturally on more than 50 islands and accidentally introduced to Guam and other islands. See implications for conservation in Rodda and Savidge (2007). (d) The Oriental Blind Snake (Indotyphlops braminus) is native to almost 60 different islands and was accidentally introduced in dozens of other islands throughout the world (Wallach 2009). This specimen was found on Boca Chica Key, Florida. (e) The Solomon Island Ground Boa (Candoia paulsoni) is perhaps the most widespread snake on islands, occurring in more than 60 islands in the Oriental region. (f) Feick’s Dwarf Boa (Tropidophis feicki) is representative of dwarf boas that occur throughout numerous islands of the Caribbean. This species occurs in Cuba, where the genus and family (Trophidophiidae) reach their highest diversity. There are 32 species of the genus Tropidophis, found in areas of South America and the West Indies, where this genus has more successfully speciated.
Sources: Photographs by Ming- Chung Tu (a), Mark O’Shea (b and e), Coleman Sheehy III (c), Jonathan Mays (d), and Javier Torres Lopez (f), all reproduced with permission.
scavenging; (6) effective means of prey acquisition; (7) infrequent feeding on relatively large prey and “slow” digestive physiology; (8) secretive behaviors and cryptic morphologies; (9) special scansorial capabilities in many species; and (10) thermal plasticity.
In this chapter, we review much of the key literature relating to islands inhabited by snakes and the insular ecology of snakes. Using a database on the occurrence of snakes on islands, we also provide broad patterns of snake diversity. Our goals are to illustrate how studies of snakes on islands can inform general principles related to biology of islands or fragmented habitat, evaluate what are considered to be novel features related to the ecology of snakes on islands that can serve to enhance understanding of complex situations, and stimulate future research as a result of forward-looking perspectives that emerge from the robust literature on snakes and islands.
Geography and Features of Islands Inhabited by Snakes
The main island types regarding their origin are oceanic islands (originated over oceanic plates), continental fragments (portions of continental rock originated by plate tectonic processes), and continental—or land-bridge— islands (located on continental shelves; Whittaker and Fernández-Palacios 2007). For practical reasons, here we combine continental fragments (e.g., Madagascar and New Caledonia) and oceanic islands (e.g., Hawaii and Canaries) into a single category, herein called oceanic islands (Pyron and Burbrink 2014; Figure 1.2). This review is concerned exclusively with islands in the sea—that is, we do not deal with lake, river, and estuary islands, including those separated from mainland by a narrow (<150 m wide) channel or similar feature. For instance, we consider Singapore as a sea island because it is separated from the remaining of the Malayan peninsula by the Strait of Johor, which is 615 m wide at its narrowest point. On the other hand, we do not consider the Cardoso Island, on the coast of southeastern Brazil, as a sea island because despite its name, the channel that separates it from mainland is only 88 m wide at its narrowest point. Finally, we do not deal with sea snakes because they generally are not confined to the islands where they occur. Additional details concerning how our database of insular snakes was built are provided in the Appendix.
The origin of snake populations in different island types differs, mainly because of variation in the origins of islands. Thus, understanding how these island types originated is crucial for understanding the diversity and biogeography of insular snakes. Land-bridge islands are originated by the variation in
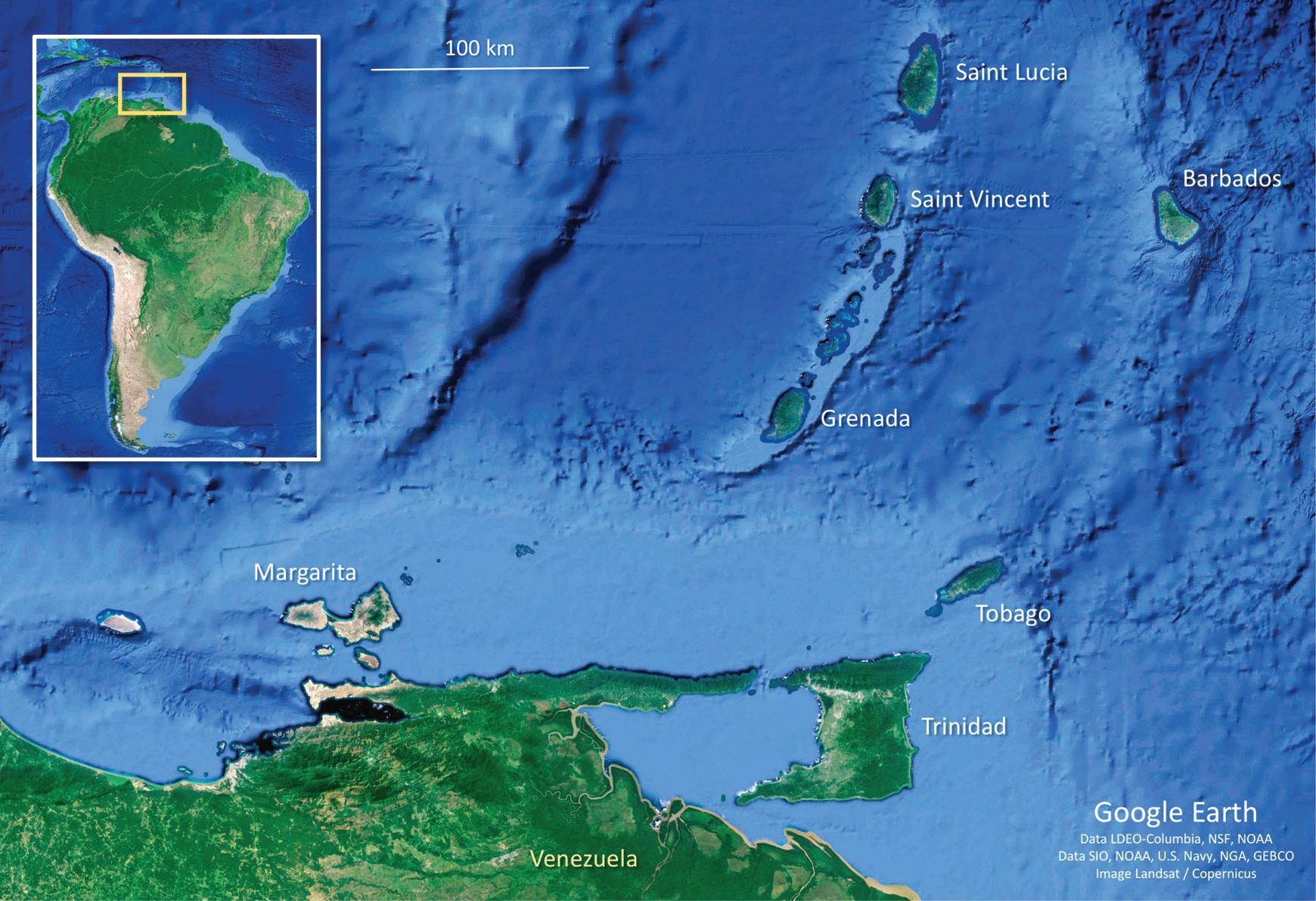
Figure 1.2 A satellite image of northern Venezuela and southern Lesser Antilles showing land-bridge (Margarita, Trinidad, and Tobago) and oceanic islands (Grenada, Saint Vincent, Saint Lucia, and Barbados). Note that land-bridge islands are located on the shallow (<100 m deep) continental shelf of coastal Venezuela, whereas oceanic islands are separated from the South American continent by deep sea (e.g., the deep strait [>700 m] that separates Grenada from the Venezuelan continental shelf).
sea level during glaciation cycles. Most of them have been connected, disconnected, and reconnected to the mainland many times during the glaciations of the past 1.8 million years (Whittaker and Fernández-Palacios 2007). For example, many islands in continental shelves throughout the world were connected to their respective continents during the last glacial maximum approximately 29,000 to 21,000 ybp, when sea level reached approximately 130 m below the present level (Lambeck et al. 2014). Extensive portions of continental shelves were exposed by then. When sea level rose to the present level, mountains amid these coastal lowlands became the land-bridge islands that we see today.
Continental fragments are originated by plate tectonics processes in which a portion of a continent moves away from the main portion, becoming an island (Whittaker and Fernández-Palacios 2007). Madagascar, for instance, was part of the Gondwana supercontinent and became separated from Africa in the Late Jurassic–Early Cretaceous and from India during the Late Cretaceous (Ali and Atchnison 2008). On the other hand, oceanic islands are originated mainly by volcanism in the areas of contact of tectonic plates,
and they were never in contact with continents, although they may be close to them in some instances. A detailed review of the origin of different island types is provided by Whittaker and Fernández-Palacios (2007).
Snakes are found on land-bridge and oceanic islands throughout the planet—on all five continents; in all oceans; and from 42 degrees south in Tasmania, Australia, to 63 degrees north at Hitra Island, Norway. Among land-bridge islands, snakes occur on islands varying in size from 1,000 m2 (small Japanese islands in the South China Sea) to approximately 786,000 km2 (New Guinea Island) and varying in elevation from less than 1 m above sea level (ASL) on Huevos Island (Trinidad and Tobago) to almost 5,000 m ASL on New Guinea Island. Among oceanic islands, those inhabited by snakes vary in area from less than 1 km2 (e.g., many islands in the Bahamas) to more than 580,000 km2 (Madagascar) and vary in elevation from less than 5 m ASL on some small islands in the Cuban Archipelago to 3,700 m ASL on Lombok Island, Lesser Sunda Islands.
Most islands inhabited by snakes are located in the Eastern Hemisphere, where most tropical and subtropical islands are concentrated (Figure 1.3). This region harbors most groups of islands on which island snakes occur, including the Japanese and Malay archipelagoes as well as the Philippines. In the Western Hemisphere, most islands inhabited by snakes are located in the Caribbean and the Bahamas. The absolute middle latitude of islands explains just a small amount (10% or less) of the variation of snake richness on both land-bridge and oceanic islands (r2 = 0.104, p < 0.001 and r2 = 0.058, p < 0.001, respectively; both variables log transformed), with islands from lower latitudes tending to harbor more species. However, using their database with islands larger than 250 km2, Pyron and Burbrink (2014) found a stronger effect (r2 = 0.25) of middle latitude on insular richness of snakes. Finally, the vegetation cover of islands inhabited by snakes includes most of the terrestrial vegetation types found on continents, from the dry Sonoran Desert on islands in the Sea of Cortez, Gulf of California, to the luxuriant rainforests of lowland Borneo.
Patterns of Diversity and Endemism
Relatively few efforts have been made to describe global patterns of insular snake diversity and to understand the processes responsible for these patterns. The few recent exceptions include Pitta et al. (2013), who explored the compositional dissimilarity of reptiles and amphibians in oceanic and land-bridge islands, and Pyron and Burbrink (2014), who explored factors that could
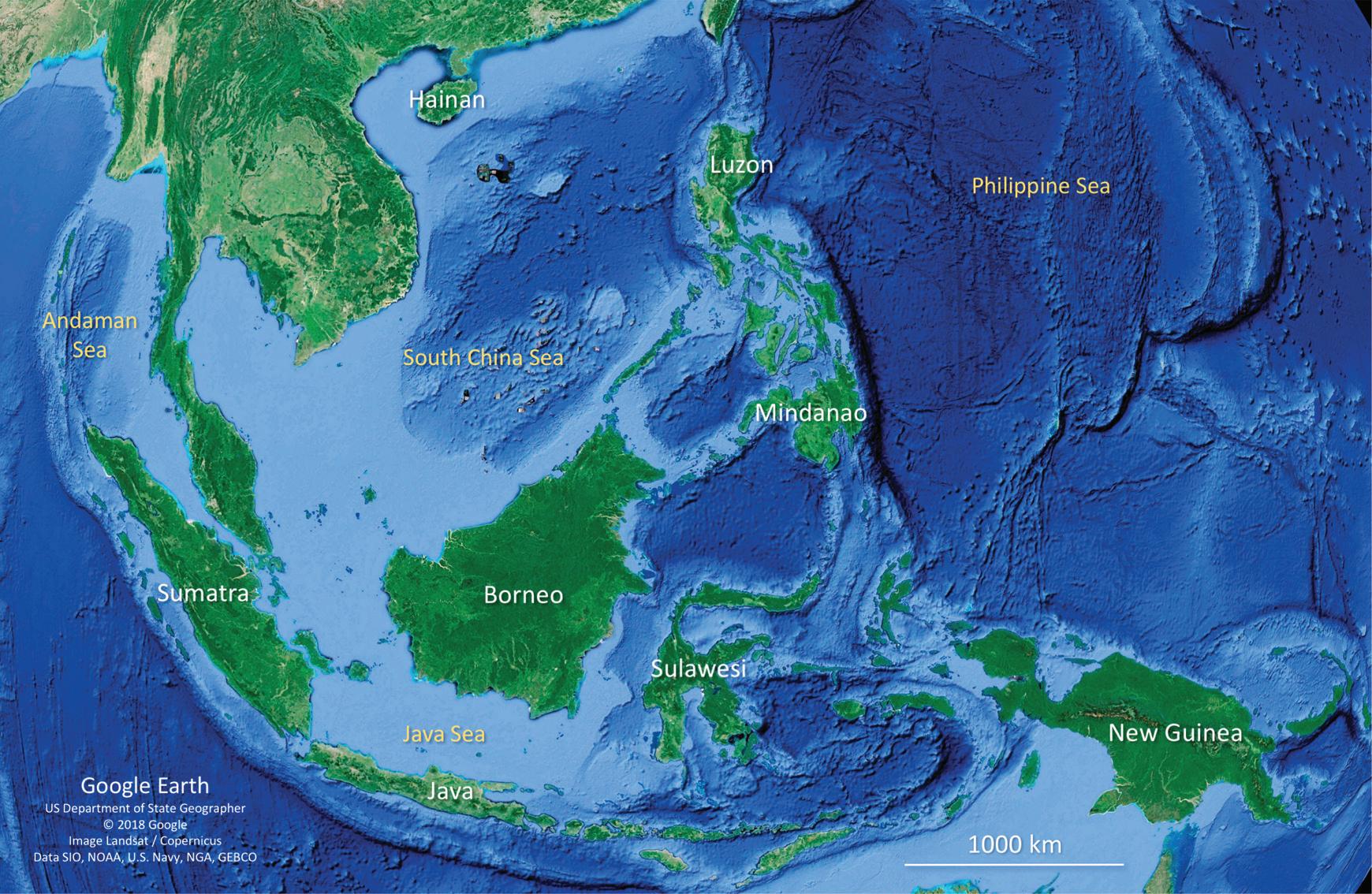
Figure 1.3 Satellite image showing a large portion of the Oriental zoogeographical region, where many of the largest and richest land-bridge islands are located, including Borneo (with 141 species of snakes), Sumatra (133 species), Java (96 species), and New Guinea (93 species). Large and rich oceanic islands are also found in this region, such as Sulawesi (with 63 species of snakes) and Luzon (51 species). Also in this region are some of the islands bearing a high number of endemic snake species, such as Sri Lanka (with 46 endemics, not shown), Borneo (31 endemics), New Guinea (26 endemics), Sumatra (19 endemics), and Sulawesi (16 endemics).
explain patterns of species richness and phylogenetic composition of snakes in oceanic islands. In this section, we provide a detailed description of the global patterns of diversity and phylogenetic composition of island snakes using a database of island snakes we built for this purpose, and we compare general patterns with those found by both Pitta et al. and Pyron and Burbrink.
Our database contains 1,223 snake species occurring on 987 islands— 368 land-bridge islands on which 718 snake species occur and 618 oceanic islands on which 761 snake species occur. Snake richness on land-bridge islands varies from 1 to 141. Although 32.6% of the land-bridge islands have only a single species and 95.4% have 20 species or less, snake richness is very high in the five largest (61,411–743,330 km2) land-bridge islands, all of them located in the Oriental and Oceanian zoogeographical regions (Holt et al. 2013; see Figure 1.3): Borneo (141 species), Sumatra (133), Java (96), New Guinea (93), and Sri Lanka (89). Among oceanic islands, snake richness varies from 1 to 89 species. Similar to land-bridge islands, 31.7% of oceanic islands have only a single species of snake and 97.1% have 20 species or
less. On the other hand, only 3 oceanic islands (with areas from 104,688 to 587,041 km2) harbor more than 50 species of snakes: Madagascar (89 species), Sulawesi (63), and Luzon (51).
The mean number of snakes per island on land-bridge islands is higher than that on oceanic islands (respectively, 6.1 ± 14.2 snakes/island; range, 1–141; N = 368; and 4.7 ± 7.6 snakes/island; range, 1–89; N = 618). Island area explains slightly more than one-third of the variation in snake richness for both island types (r2 = 0.367, p < 0.001 for land-bridge islands; r2 = 0.375, p < 0.001 for oceanic islands; both variables log transformed). Fattorinni (2010) failed to find an effect of island size on lizard and snake richness on Italian land-bridge islands. On the other hand, using Bayesian models, Pyron and Burbrink (2014) found a significant effect of island size (as well as temperature, isolation, and elevation) on snake richness on oceanic islands.
The diversity of habitats on an island also may be a good predictor of species richness, probably because more niches are available on more heterogeneous islands (MacArthur and Wilson 1967; Ricklefs and Lovette 1999; Whittaker and Fernández-Palacios 2007). In general, larger islands typically show higher habitat heterogeneity compared to small islands (MacArthur and Wilson 1967; Whittaker and Fernández-Palacios 2007). Because habitat diversity tends to be strongly correlated with island maximum elevation (Ricklefs and Lovette 1999; Henderson 2004; Kalmar and Currie 2007), we used the maximum elevation of islands as a proxy of habitat heterogeneity ( cf . Pyron and Burbrink 2014) to explore the effect of habitat diversity on snake richness using our database of island snakes. The maximum elevation of islands (both types) explains onethird of the variation in snake richness ( r 2 = 0.326, p < 0.001), indicating that habitat heterogeneity explains at least part of the variation of snake richness on islands. When island types are considered separately, maximum elevation explains more variation of snake richness on land-bridge islands than on oceanic islands ( r 2 = 0.403, p < 0.001 and r 2 = 0.292, p < 0.001, respectively). Henderson (2004) also provides evidence for an effect of habitat diversity on snake richness on the Lesser Antillean islands.
Compared to similar-sized regions of the mainland, islands tend to show higher levels of endemism (Kier et al. 2009). Approximately one-tenth (95) of the 987 islands in our database have at least one endemic snake (i.e., a species that occurs on that island and nowhere else). Endemics are found on both land-bridge and oceanic islands, and the proportion of islands harboring endemics is higher on oceanic (11.6%) than on land-bridge islands (6.2%).
However, the mean number of endemic snakes on land-bridge islands (6.9 ± 12.0; range, 1– 46; N = 23) is double that on oceanic islands (3.5 ± 9.1; range, 1–72; N = 72). Among land-bridge islands, the number of endemics is especially high in Sri Lanka (46 endemics), Borneo (31), New Guinea (26), and Sumatra (19). Among oceanic islands, the number of endemics is especially high in Madagascar (71 endemics), Cuba (28), Hispaniola (17), and Sulawesi (16). When endemicity is considered in relation to island size, land-bridge islands have on average a much higher number of endemics per island area (0.68 ± 1.54 endemics/km2; range, <0.01 to 6.67 endemics/km2; N = 23) compared to oceanic islands (0.26 ± 1.36 endemics/km2; range, <0.01 to 11.11 endemics/km2; N = 72). Highest endemicity per area of island is found on very small land-bridge islands (Franceses and Queimada Grande Islands off the coast of eastern Brazil, with 0.2 and 0.4 km2, respectively) and oceanic islands (Great Bird Island in the Lesser Antilles and Round Island in the Mascarenes, with 2.15 and 0.09 km2, respectively).
Among the 1,223 insular snakes in our database, 718 occur on land-bridge islands, 776 on oceanic islands, and 249 on both land-bridge and oceanic islands. Insular snakes occur on 1– 48 land-bridge islands (3.1 ± 5.1 islands, N = 718) and on 1–59 oceanic islands (3.9 ± 6.4 islands, N = 761). Most snakes from land-bridge islands occur on few islands: Slightly more than half (55.7%) occur on a single island, 13.6% occur on two islands, and 87.3% occur on five or less islands. Similar proportions are found in snakes from oceanic islands: Approximately half (50.0%) occur on a single island, 15.0% occur on two islands, and 83.2% occur on five or less islands. At the other extreme, 50 species (4.1%) occur on more than 20 islands (both island types), of which there are 29 colubrids, 5 boids, 5 typhlopids, 4 viperids, 2 lamprophiids, 2 pythonids, and 1 species of each of the families Acrochordidae, Elapidae, and Homalopsidae. Most of these species are from the Oriental and/or Oceanian zoogeographical regions. Only 7 species occur on 50 or more islands (both types): Telescopus fallax (Colubridae, 50 islands), Boiga irregularis (Colubridae, 54 islands), Malayopython reticulatus (Pythonidae, 55 islands), Natrix natrix (Colubridae, 55 islands), Indotyphlops braminus (Typhlopidae, 57 islands), Cerberus rynchops (Homalopsidae, 60 islands), and Candoia paulsoni (Boidae, 61 islands) (see Figure 1.1).
The composition of insular faunas is highly variable and strongly dependent on the ability to disperse overseas (Whittaker and Fernández-Palacios 2007). For groups with high dispersal ability, faunal composition tends to be similar in nearby islands, whereas groups with lower ability to disperse overseas tend to show a higher dissimilarity of composition among islands. Pitta et al.