Cosmetic Dermatology Fellow, Department of Dermatology and Cutaneous Surgery, Miller School of Medicine, University of Miami, Miami, Florida; Dermatologic Clinic, University Hospital of Zurich Zurich, Switzerland
Chapters 33 and 34
K.P.Ananth Chapter 31
Nidhi J.Avashia,BS
Miller School of Medicine, University of Miami, Miami, Florida Chapter 29
Marianna L.Blyumin,MD
Dermatology Resident, Department of Dermatology and Cutaneous Surgery, Miller School of Medicine, University of Miami, Miami, Florida Chapter 23
Angela S.Casey,MD
Assistant Professor, Dermatology and Mohs Surgery, University of Vermont College of Medicine, Fletcher Allen Health Care, Burlington, Vermont Chapter 26
Maria Paz Castanedo-Tardan,MD
Department of Dermatology and Cutaneous Surgery, Miller School of Medicine, University of Miami Miami, Florida
Chapters 29, 35, 38, and 39
CONTRIBUTORS
Mohamed L.Elsaie,MD,MBA
Cosmetic Dermatology Fellow, Department of Dermatology and Cutaneous Surgery, Miller School of Medicine, University of Miami, Miami, Florida; Department of Dermatology and Venereology, National Research Center, Cairo, Egypt
Chapters 10 and 22
Lisa Danielle Grunebaum,MD
Assistant Professor, Division of Facial Plastic and Reconstructive Surgery, Department of Otolaryngology and Head and Neck Surgery, University of Miami, Miami, Florida
Chapter 22
Sharon E.Jacob,MD
Assistant Professor, Divisions of Medicine and Pediatrics (Dermatology), University of California, San Diego, San Diego, California
Chapter 18
H.Ray Jalian,MD
Resident Physician, Department of Medicine, Division of Dermatology, David Geffen School of Medicine at UCLA, Los Angeles, California
Chapter 4
Joely Kaufman,MD
Assistant Professor, Department of Dermatology and Cutaneous Surgery and Director of Laser and Light Therapy, University of Miamia Cosmetic Medicine and Research Institute, Miami, Florida
Chapter 24
Jonette Keri,MD,PhD
Assistant Professor, Miller School of Medicine, University of Miami, Miami, Florida; Chief, Dermatology Service, Miami VA Hospital, Miami, Florida
Chapters 15 and 16
Jenny Kim,MD,PhD
Associate Professor, Department of Medicine and Division of Dermatology, David Geffen School of Medicine at UCLA, Los Angeles, California
Chapter 4
Suzan Obagi,MD
Assistant Professor of Dermatology, Director, The Cosmetic Surgery and Skin Health Center, University of Pittsburgh Medical Center, Pittsburgh, Pennsylvania
Chapters 3 and 26
Sogol Saghari,MD
Department of Dermatology, University of Miami, Miami, Florida; Private Practice, Los Angeles, California
Chapters 1, 2, 7, 13, 16, 19, 20, 21, 23, and 30
Susan Schaffer,RN
University of Miami, Cosmetic Medicine and Research Institute, Miami Beach, Florida
Chapter 21
Stuart Daniel Shanler,MD,FACMS
Private Practice, New York, New York
Chapter 16
Anita Singh,MS
Miller School of Medicine, University of Miami, Miami, Florida
Chapter 3
Kumar Subramanyan,PhD
Senior Manager, Consumer and Clinical Evaluation, Unilever Global Skin Research & Development Shanghai, China
Chapter 31
Voraphol Vejjabhinanta,MD
Postdoctoral Fellow, Mohs, Laser and, Dermatologic Surgery, Department of Dermatology and Cutaneous Surgery, Miller School of Medicine,University of Miami, Miami, Florida; Clinical Instructor Suphannahong Dermatology Institute, Bangkok, Thailand Chapter 3
Edmund Weisberg,MS
Managing Editor, Center for Clinical Epidemiology and Biostatistics, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania Chapters 9, 28, 36, 37, and 40
Heather Woolery-Lloyd,MD
Assistant Professor, Department of Dermatology and Cutaneous Surgery, Director of Ethnic Skin Care University of Miami Cosmetic Medicine and Research Institute, Miami, Florida Chapter 14
Larissa Zaulyanov-Scanlan,MD
Voluntary Faculty, University of Miami Cosmetic Medicine and Research Institute, Miami Beach, Florida; Private Practice, Delray Beach, Florida Chapters 5 and 25
PREFACE
Cosmetic dermatology is a rapidly growing field that can attribute its popularity to aging baby boomers. Although many dermatologists perform cosmetic procedures and millions of dollars are spent each year on cosmetic products, there is a paucity of published research inthis field. I was stimulated to write this text because I have found it challenging to conduct thorough research in preparation for my lectures and articles on cosmetic science as there exists no undisputed reference at the moment. Of the research performed by cosmetic scientists, much of it, unfortunately, is proprietary information owned by corporations and is not published or shared in any way for the immediate benefit of the medical community and other cosmetic professionals. This results in each company or cosmetic scientist having to “reinvent the wheel.” My goal, with this book, is to create a link, featuring a better streaming flow of information, between the fields of dermatology and cosmetic science. This text is designed to help cosmetic dermatologists understand the available information on various cosmetic products and procedures. It should also help cosmetic chemists to understand the issues that cosmetic dermatologists deal with on a frequent basis. In addition, this text should fill the gap in knowledge among professionals such as aestheticians who need to know what to apply to patients’ or clients’ skin and about the products that people purchase over-the-counter and apply to their skin. This text should help these professionals answer the questions that their clients/patients ask about skin care products and their scientific validity. It is my hope that this text will encourage cosmetic dermatologists, cosmetic scientists and aestheticians to insist upon well researched cosmetic products
and procedures. By working together in this way we can preserve the integrity of an exciting and rapidly developing field of study.
Research in the field of cosmetic dermatology should be encouraged for many reasons. Obviously, it is vital to maintain the hard earned integrity of the field of dermatology. In addition, the discoveries made though cosmetic dermatology research will likely benefit other fields of dermatology. For example, research into the anti-aging effects of antioxidants may lead to enhanced knowledge of chemopreventive techniques to be used to prevent skin cancer. Advances in acne therapy, vitiligo and other disorders of pigmentation are also possible. In fact, it is interesting to note that the development of Vaniqa™, a cream designed to slow hair growth in women with facial hair, has led to the availability of an intravenous treatment for African Sleeping Sickness, a major cause of death in Africa. Without the financial incentive to develop Vaniqa, which is used for purely aesthetic purposes, this life-saving drug would not be available. For many reasons, all pharmaceutical, medical device, and cosmetic companies should be encouraged to research their products.
Although there is much research performed by cosmetic companies on the effects of cosmetics on the skin, much of this data is proprietary and is not published nor shared with the rest of the scientific community. The reasons for this are numerous, but competition between companies and the desire to be the first to come out with a new “miracle product” are prominent among them. However, the issue is even more complex. The FDA has different definitions for drugs and cosmetics. Cosmetic prod-
ucts do not have to be researched in any standard way because FDA approval is not required. Instead, cosmetic products are voluntarily registered by the companies that develop them. However, drugs must undergo years of expensive trials establishing both safety and efficacy before receiving FDA approval (see Ch 28). This disparity means that a company is more reluctant to publish data that could cause their product to be labeled as a drug.
The dearth of published data on cosmetic products has forced physicians, aestheticians, and lay people to rely on sales people and marketing departments to obtain information about cosmetic formulations. This has led to much misinformation that has diminished the credibility of cosmetic products and the cosmetic field in general. Because an ever-increasing number of dermatologists and other physicians are practicing “cosmetic dermatology,” it is imperative that the cosmetic dermatologist practice evidence-based medicine in order to distinguish efficacious treatments from mere marketing hype. This text sifts through the knowledge of the effects cosmetic products and procedures have on the skin and its appearance. The amount of research that should still be performed is daunting; however, the field is young and the rewards are great. I encourage everyone to join me in the exciting endeavor to find scientifically proven methods of improving the appearance of the skin.
Leslie Baumann, MD
“Don’t worry if your job is small, And your rewards are few. Remember that the mighty oak, Was once a nut like you.”
Anonymous
This page intentionally left blank
The first edition of this book was printed in 4 languages and was the bestselling textbook on cosmetic dermatology worldwide (or so I have been told). There are many people to thank for this and the many wonderful things that have occurred in the last 6 years. First I would like to thank Dr. Stephen Mandy who took me in as a newly graduated resident in 1997, and let me and my husband live with him for two weeks while he taught me about the newly emerging field of cosmetic dermatology. (I learned to inject collagen on his secretary!) That was the beginning of what has now been an 11-year friendship. Dr. Francisco Kerdel negotiated my first job and office space and he and Dr. William Eaglstein mentor me to this day. They were thanked in the first edition but I will never be able to thank them enough for what they have done for me.
This year, the University of Miami Miller School of Medicine decided to create the Cosmetic Medicine and Research Institute (CMRI), which consists of cosmetic dermatology, oculoplastic surgery, facial plastic surgery and nutrition. The role of this multi-specialty institute is to provide cutting edge dermatologic and surgical procedures to enhance appearance. By combining accomplished physicians from the various cosmetic specialties, the Institute can offer patients the expertise of many different types of physicians in order to achieve the best outcome. The mission of the Institute is to perform research in the area of cosmetic medicine, and many genetic initiatives to look for the genetic influences on appearance have begun. In addition, the CMRI will provide training to physicians on cosmetic dermatology and cosmetic procedures.(See www.derm.net for more information.)
ACKNOWLEDGMENTS
I am very proud to announce that I have been selected to be the Director of the University of Miami Cosmetic Medicine and Research Institute. For this honor I would like to thank several people for believing in me and giving me this opportunity:
Pascal Goldschmidt, MD (the Dean of the University of Miami Medical School) – Dr. Goldschmidt is a true visionary and a leader in the field of the genetic influences in atherosclerosis. He opened the doors to basic science research for me and shared his genetic research team with me until I could find funding. In addition, he did the great honor of introducing me to Bart Chernow, MD and William O’Neil, MD (both of whom are Vice Deans at the University of Miami). The three of them appointed me Director of the University of Miami Cosmetic Medicine and Research Institute and gave me one of the most wonderful opportunities of my life. Dr. Chernow is a brilliant man and a true magician because he can pull all kinds of opportunities and ideas and innovations “out of his hat.” I consider Bart and his wife Peggy good friends and I thank them both for their support.
I would like to thank David Seo, MD, my partner on the genetic trials, for his patience in getting me up to speed on genetic research. My fingers are crossed that we will discover great things together in the next 2 years. Thanks to the doctors who are a part of the CMRI and have chapters in this text. They have all taught me so much and are great to work with: Drs. Lisa Grunebaum, Joely Kaufman, Wendy Lee, Heather Woolery-Lloyd,
and Larissa Zaulyanov-Scanlan. Thanks to Neal Shapiro for handling the financial aspect of the Institute so that I can concentrate on my true loves…seeing patients and performing research. Huge hugs and thanks to Susan Schaffer-RN who is my great friend, confidant, and Head of Nursing for the CMRI. She travels around the world with me, lecturing on cosmetic issues and helping to keep me sane. Edmund Weisbergyou are hilarious and fun to work with. I would never have written the first edition of this book without you! Stephanie and Fransheley- you have worked with me for many years and I have loved it and I look forward to MANY more.
I would like to thank Catherine Drayton and Richard Pine, my book agents for my NY Times bestselling book called “The Skin Type Solution” (Bantam 2005) (www.skintypesolutions.com). They negotiated an unprecedented book deal for me and are the best in the field. I first unveiled the Baumann Skin Typing System in this book. Catherine- Thanks for all the attention that you give to me in spite of the fact that we live on opposite sides of the world (and thanks for taking me sailing with you in Australia when I was there for the book launchthat was SO COOL!). I will never forget the support that Irwin Applebaum and his amazing team at Bantam Dell (a division of Random House) gave The Skin Type Solution when it launched. Phillip Rappaport is a great editor and friend.
I would like to thank my family, to whom this book is dedicated. My husband Roger and my sons Robert
and Max are a constant source of joy and strength for me. I love cooking with them! I am fortunate to be very close with both my mother, Lynn McClendon, and my mother-in-law, Josie Kenin. They are great role models and friends and I am very lucky to have them. Thanks to my friends Jill Cooper, Melina Goldstein, Sofie Matz and Debbie Kramer for listening to me and keeping me calm.
Dr. Sogol Saghari, who was my fellow for one year and now has a dermatology practice in Los Angeles, made huge contributions to this book. She helped on the first draft of
many of the chapters. She is a brilliant dermatologist and an incredibly nice person. I was so lucky to have her as a fellow. Thanks to all the doctors who contributed to the chapters in this book. Special thanks to Mohammed Lotfy, MD, who was available 24 hours a day helping me with literature searches and drawing the illustrations. He is one of the most dedicated dermatologists I have ever met. Inja Bogdan, MD and Maria Paz Castanedo-Tardan, MD were also fellows that contributed chapters and have great careers ahead of them. And last but certainly not least-
I would like to thank Anne Sydor for convincing me to write the second edition of this book. I never would have been able to get up at 5am and get this done if you had not encouraged me. Thanks for being my cheerleader and for lighting a fire in me to get this done ... FINALLY! I am so proud of this book and poured my soul into it. I hope that all of you enjoy reading it as much as I enjoyed writing it.
Affectionately,
Leslie Baumann, MD
1 SECTION
Basic Concepts of Skin Science
This page intentionally left blank
CHAPTER 1
Basic Science of the Epidermis
Leslie Baumann,MD
Sogol Saghari,MD
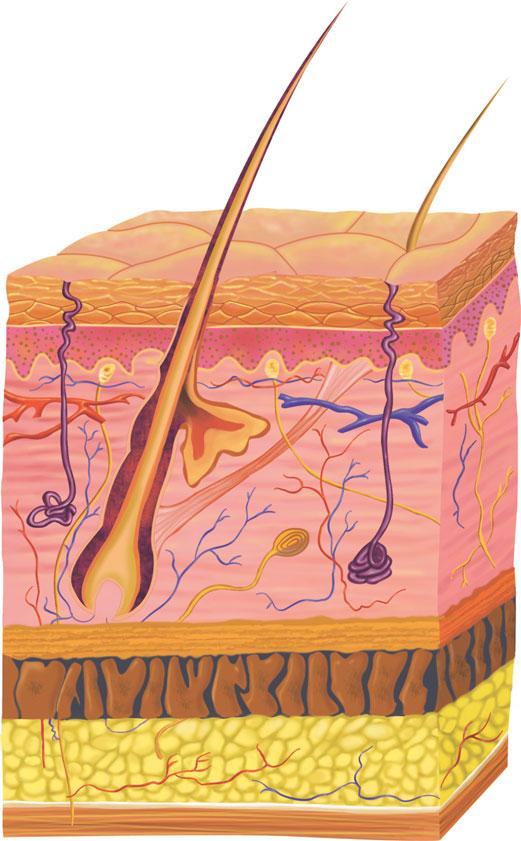
The skin is composed of three primary layers: epidermis, dermis, and subcutaneous tissue. Each layer possesses specific characteristics and functions. Although research regarding skin layers continues, much is already known about the structure of each component. New discoveries about these components have already led to prenatal diagnoses of many inherited diseases and to improved therapies. In the future, study of these components will likely lead to an enhanced understanding of skin aging and the effects of topical products on the biologic function of the skin. The epidermis is the most superficial layer of the skin. It is very important from a cosmetic standpoint, because it is this layer that gives the skin its texture and moisture, and contributes to skin color. If the surface of the epidermis is dry or rough, the skin appears aged. Knowledge of the basic structure of the epidermis best enables a practitioner to improve the appearance of patients’ skin.
THE KERATINOCYTE
Keratinocytes, also known as corneocytes, are the cells that comprise the majority of the epidermis. Keratin filaments are major components of the keratinocytes, and provide structural support. There are two types of keratin filaments: acidic (type I, K10–20) and basic (type II, K 1–10). They both must be expressed for a keratin filament to
develop.1 In other words, an acidic type and a basic type are always expressed together and they form a keratin filament together. Keratinocytes are “born” at the base of the epidermis at the dermal–epidermal junction (DEJ). They are produced by stem cells, which are also called basal cells because they reside at the base, basal layer, of the epidermis. When the stem cells divide, they create “daughter cells,” which slowly migrate to the top of the epidermis. This process of daughter cells maturing and moving to the top is called keratinization.
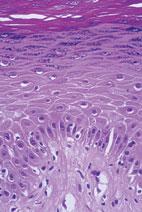
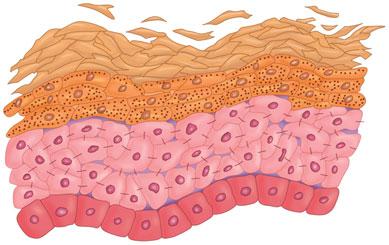
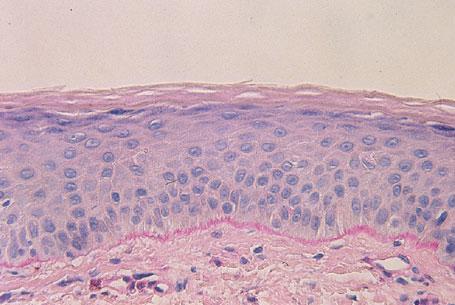
As these cells progress through the epidermis and mature, they develop different characteristics. The layers of the epidermis are named for these characteristic traits. For example, as mentioned, the first layer is the basal layer because it is located at the base of the epidermis. Basal cells are cuboidal in shape. The next layer is referred to as the spinous layer because the cells in this layer have prominent, spiny attachments called desmosomes. Desmosomes are complex structures composed of adhesion molecules and other proteins and are integral in cell adhesion and cell transport. The next layer is the granular layer, named so because these cells contain visible keratohyaline granules. The last, outermost layer is the stratum corneum (SC), a condensed mass of cells that have lost their nuclei and granules (Figs. 1-1 and Fig. 1-2). The SC is covered by a protein material called the cell envelope, which aids in providing a barrier to water loss and absorption of unwanted materials.
As keratinocytes migrate through the layers of the epidermis, their contents and functions change according to, or depending on, the specific epidermal layer in which they are moving. Although the functions of the keratinocyte have not been completely elucidated, many of them are understood. It is known
Keratohyaline granules
Desmosomes
Stratum corneum
Granular layer
Spinous layer
Basal layer
Dermis
FIGURE 1-2 Histopathology of the epidermis demonstrating the four layers.(Image courtesy of George Ioannides,MD.)
that keratinocyte activity, such as the release of cytokines, can be affected by topical products administered to the skin. Keratinocytes and their components at each level of the epidermis starting at the basal layer and proceeding to the superficial layers of the epidermis are described below.
Keratinocyte Function
THE BASAL LAYER (STRATUM BASALE) Basalcells join with other basal and the overlying spinous cells via desmosomes, thus forming the basement membrane. These basal keratinocytes contain keratins 5 and 14, mutations in which result in an inherited disease called epidermolysis bullosa simplex. Keratins 5 and 14 are presumed to establish a cytoskeleton that permits flexibility of the cells. This flexibility allows cells to proceed out of the basal layer and migrate superficially, thus undergoing the keratinization process.
Keratohyaline granule
Desmosome
Stratum corneum
Granular layer
Spinous layer
Basal layer
Basal cells are responsible for maintaining the epidermis by continually renewing the cell population. Of the basal layer, 10% of cells are stem cells, 50% are amplifying cells, and 40% are postmitotic cells. Normally, stem cells are slowly dividing cells, but under certain conditions such as wound healing or exposure to growth factors, they divide faster. They give rise to transient amplifying cells. Transient amplifying cells are responsible for most of the cell division in the basal layer and produce postmitotic cells, which undergo terminal differentiation and move superficially to become suprabasal cells that continue their upward migration to become granular cells and ultimately part of the SC (Fig. 1-3)
THE SPINOUS LAYER (STRATUM SPINOSUM)
Keratins 1 and 10 are first seen in this layer of suprabasal keratinocytes. These keratins form a more rigid cytoskeleton
FIGURE 1-1 The layers of the epidermis.
DERMIS
Desquamating cell
Stem Cells
FIGURE 1-3 The stem cells divide and produce amplifying cells that greatly increase the number of keratinocytes.These in turn become the mature,terminal,and differentiated cells.The numbers indicate the cell generation.
that confers greater mechanical strength to the cell. It is worth mentioning that under hyperproliferative conditions such as actinic keratosis, wound healing, and psoriasis, keratins 6 and 16 are upregulated in the suprabasal keratinocytes.
Lamellar granules, which are considered the first sign of keratinization, first appear in this layer. They contain lipids such as ceramides, cholesterol, and fatty acids as well as enzymes such as proteases, acid phosphatase, lipases, and glycosidases. It has been recently shown that cathelicidin, an antimicrobial peptide, is also stored in the lamellar granules.2 These granules migrate to the surface and expel their contents by exocytosis. The released lipids coat the surface, imparting barrier-like properties. Desmosomes are very prominent in this layer, thus accounting for the name “spinous layer.”
The advanced stage of differentiation of suprabasal keratinocytes is conducive to staining for products not found on basal cells (i.e., sugar complexes and blood group antigens). The cytoplasm contains proteins not found in the lower layers such as involucrin, keratolinin, and loricrin. These proteins become cross-linked in the SC to confer strength to the layer.
THE GRANULAR LAYER (STRATUM GRANULOSUM) Granular layer keratinocytes reside in the uppermost viable layer of the epi-
dermis. The “granules” represent keratohyaline granules, which contain profilaggrin, the precursor to filaggrin. The protein filaggrin cross-links keratin filaments providing strength and structure. The proteins of the cornified cell envelope (involucrin, keratolinin, pancornulins, and loricrin) are cross-linked in this layer by the calcium-requiring enzyme transglutaminase (TGase) to form the cell envelope. There are four types of transglutaminases present in the epidermis: TGase 1 or keratinocyte TGase, TGase 2 or tissue TGase, TGase 3 or epidermal TGase, and TGase 5. Only TGases 1, 3, and 5 participate in the development of the corneocyte envelope (CE) formation. TGase 2 has other functions including a role in apoptosis (programmed cell death). It is known that TGase activity increases with
BOX 1-1
the elevation of Ca2+ levels in the medium of cultured keratinocytes.3 This in turn results in the formation of the cornified cell envelope and differentiation of keratinocytes.4,5 The active metabolite of vitamin D, known as 1,25dihydroxyvitamin D3 [1,25(OH)2D3], also plays a role in keratinocyte differentiation (Box 1-1). It enhances the Ca2+ effect on the keratinocytes, and increases transglutaminase activity as well as involucrin levels,6 the combined effects of which induce CE formation.7,8
Calcium is known to be an inducer of differentiation and a suppressor of proliferation in epidermal keratinocytes.9,10 It has been shown that in the state of low Ca2+ levels (0.05 mM), keratinocytes are in a proliferative stage, while increases in Ca2+ levels (0.10–0.16 mM) lead to expression of differentiation markers such as keratins 1 and 10, TGase, and filaggrin.9
Granular cells exhibit anabolic properties such as synthesis of filaggrin, cornified cell envelope proteins, and high molecular weight keratins. In addition, they show catabolic events such as dissolution of the nucleus and organelles.
THE HORNY LAYER (SC) The most superficial layer of the epidermis is the SC or horny layer, which is, on average, approximately 15-cell layers thick.13,14 The keratinocytes that reside in this layer are the most mature and have completed the keratinization process. These keratinocytes contain no organelles and their arrangement resembles a brick wall. The SC is composed of protein-rich corneocytes embedded in a bilayer lipid matrix assembled in a “brick and mortar” fashion. The “bricks” are composed of keratinocytes and the “mortar” is made up of the contents extruded from the lamellar granules including lipids and proteins (Fig. 1-4). Cells of the midcornified layer have the most amino acid content and therefore have the highest capability for binding to water, while the
1,25-Dihydroxyvitamin D3 [1,25(OH)2D3] stimulates differentiation and prohibits proliferation of the keratinocytes.It exerts its effects via the nuclear hormone receptor known as vitamin D receptor (VDR).VDR operates with the aid of coactivator complexes.There are two known coactivator complexes:vitamin D interacting protein complex (DRIP) and the p160 steroid receptor coactivator family (SRC/p160).It has been proposed that the DRIP mediator complex is involved in proliferation and early differentiation while the SRC/p160 complex is engaged in advanced differentiation.11 The vitamin D receptors of undifferentiated keratinocytes bind to the DRIP complex,inducing early differentiation markers of K1 and K10. 12 The DRIP complex on the vitamin D receptor is then replaced by the SRC complex.The SRC complex induces gene transcription for advanced differentiation,which occurs with filaggrin and loricrin. 12 The replacement of the DRIP complex with the SRC complex on the vitamin D receptor is believed to be necessary for keratinocyte differentiation.It is important to realize that vitamin D levels are lower in older people and that this reduction may play a role in the slower wound healing characteristic in the elderly.
Intercellular lipids(fats)
Desmosomes
Keratinocytes
FIGURE 1-4 The desmosomes form attachments between the keratinocytes.The keratinocytes are surrounded by lipids.These structures form the skin barrier.
deeper layers have less water-binding capacity.15 The SC is described as the “dead layer” of cells because these cells do not exhibit protein synthesis and are unresponsive to cellular signaling.16
The horny layer functions as a protective barrier. One of its protective functions is to prevent transepidermal water loss (TEWL). Amino acids and their metabolites, which are by-products formed from the breakdown of filaggrin, comprise a substance known as the natural moisturizing factor (NMF). Intracellularly-located NMF and lipids released by the lamellar granules, located extracellularly, play an important role in skin hydration, suppleness, and flexibility (see Chapter 11).
The Cell Cycle
The above keratinization process is also referred to as the “cell cycle.” The normal cell cycle of the epidermis is from 26 to 42 days.17 This series of events, known also as desquamation, normally occurs invisibly with shedding of individual cells or small clumps of cells. Disturbances of this process may result in the accumulation of partially detached keratinocytes, which cause the clinical findings of dry skin. Disease states may also alter the cell cycle. For example, psoriasis causes a dramatic shortening of the cell cycle, resulting in the formation of crusty cutaneous eruptions. The cell cycle lengthens in time as humans age.18 This means that the cells at the superficial layer of the SC are older and their function may be impaired. Results from such compromised functioning include slower wound healing and a skin appearance that is dull and lifeless. Many cosmetic products such as retinol and alpha hydroxy acids are believed to
quicken the pace of the cell cycle, yielding younger keratinocytes at the superficial layers of the SC, thus imparting a more youthful appearance to the skin.
GROWTH FACTORS
Growth factors can be classified into two groups: proliferative and differentiative factors. Proliferative factors engender more DNA synthesis and result in proliferation of the cells. Differentiative factors inhibit the production of DNA and suppress growth, thereby resulting in differentiation of the keratinocytes. Epidermal growth factor (EGF) is one of the integral chemokines in the regulation of growth in human cells. It binds to the epidermal growth factor receptor (EGFR) located on the basal and suprabasal cells in the epidermis and activates tyrosine kinase activity, which ultimately results in proliferation of the cells.19 Keratinocyte growth factor (KGF), a member of the fibroblast growth factor family, also has a proliferative effect via the tyrosine kinase receptor on epidermal cells.20 It has been shown that KGF contributes to and enhances wound healing.21 In addition, KGF has been demonstrated to enhance hyaluronan synthesis in the keratinocytes.22 Other important growth factors include the polypeptide transforming growth factors, which consist of two types: Transforming growth factor alpha (TGF- ) and transforming growth factor beta (TGF- ). They differ in both configuration and function. TGF- is a proliferative factor, similar to EGF, and works by stimulating a tyrosine kinase response. TGF- , which includes three subtypes (1–3), is a differentiative factor with a serine/
threonine kinase receptor. TGF- 1 and TGF- 2 are present in small amounts in the keratinocytes. The presence of calcium, phorbol esters, as well as TGF- itself increases the epidermal TGF- level and promotes differentiation.23 TGF- has also been proven to have a role in scarring, and antibodies to this factor have been shown to decrease the inflammatory response in wounds and reduce scarring.24, 25
ANTIMICROBIAL PEPTIDES
Antimicrobial peptides (AMPs) have recently become an area of interest because of their involvement in the innate immune system of human skin. AMPs exhibit broad-spectrum activity against bacteria, viruses, and fungi.26,27 The cationic peptide of the AMPs attracts the negatively charged bacteria, becoming pervasive in the bacterial membrane in the process, and ultimately eliminates the bacteria. Cathelicidin and defensin are the two major groups of AMPs believed to have an influence in the antimicrobial defense of the skin. Cathelicidin has been identified in the keratinocytes of human skin at the area of inflammation, as well as in eccrine and salivary glands.28–30 In addition to antimicrobial activity, cathelicidin LL-37 demonstrates a stimulatory effect on keratinocyte proliferation in the process of wound healing.31 Pig cathelicidin PR-39 has been shown to induce proteoglycans production (specifically, syndecan-1 and -4) in the extracellular matrix in wound repair.32 Defensin is also expressed in the human keratinocytes33 and mucous membranes.34,35 -Defensin 1 seems to promote differentiation in the keratinocytes by increasing expression of keratin 10.36 Interestingly, UVB radiation has been shown to increase the levels of human -defensin mRNA in the keratinocytes.37
AMPs have been demonstrated to be involved in several dermatologic conditions including atopic dermatitis, psoriasis, and leprosy,27 as well as wound healing, all of which are beyond the scope of our discussion. The role of AMPs in the epidermal barrier will be discussed in Chapter 11.
MOISTURIZATION OF THE SC
The main function of the SC is to prevent TEWL and regulate the water balance in the skin. The two major components that allow the SC to perform this role are lipids and the NMF.
BOX 1-2
Filaggrin,named for filament aggregating protein,derived its name from the fact that it binds keratin filaments to form a structural matrix in the SC.Genetic defects in the filaggrin gene are known to play a role in a subset of ichthyosis vulgaris cases.38 Interestingly,filaggrin is not present in the superficial layers of the SC.Studies have shown that it is completely degraded into amino acids within 2 to 3 days of profilaggrin formation and its constituents are further metabolized to form the NMF.40 This is nature’s way of keeping its water-binding capabilities in the top layer of the SC where they are needed while preventing the lower layers of the SC from being disrupted by having too much water present.In addition, the level of NMF is regulated by the water activity present in the SC.
Natural Moisturizing Factor
Released by the lamellar granules, NMF is composed of amino acids and their metabolites, which are by-products formed from the breakdown of filaggrin (Box 1-2). NMF is found exclusively inside the cells of the SC and gives the SC its humectant (water-binding) qualities (Fig. 1-5). NMF is composed of very water-soluble chemicals; therefore, it can absorb large amounts of water, even when humidity levels are low. This allows the SC to retain a high water content even in a dry environment. The NMF also provides an important aqueous environment for enzymes that
require such conditions to function. The importance of NMF is clear when one notes that ichthyosis vulgaris patients, who have been shown to lack NMF, manifest severe dryness, and scaling of the skin.38 It has been demonstrated that normal skin exposed to normal soap washing has significantly lower levels of NMF when compared to normal skin not washed with surfactants.39 NMF levels have also been reported to decline with age, which may contribute to the increased incidence of dry skin in the elderly population (see Chapter 11).
Lipids
In order of abundance, the composition of skin surface lipids includes triglycerides, fatty acids, squalene, wax esters, diglycerides, cholesterol esters, and cholesterol.41 These lipids are an integral part of the epidermis and are involved in preventing TEWL and the entry of harmful bacteria. They also help prevent the skin from absorbing water-soluble agents. For decades it has been known that the absence of lipids in the diet leads to unhealthy skin (see Chapter 11). More recently, it has been shown that inherited defects in lipid metabolism, such as the deficiency of steroid sulfatase seen in Xlinked ichthyosis, will lead to abnormal skin keratinization and hydration.42 It is now known that SC lipids are affected by age, genetics, seasonal variation, and diet. Deficiency of these lipids predisposes the individual to dry skin. This has been
Intercellular lipids (mortar)
demonstrated in mice with essential fatty acid deficiency (EFAD); when fed a diet deficient in linoleic acid these mice developed increased TEWL.43 Interestingly, administration of hypocholesterolemic drugs has also been associated with dry skin changes.44
Skin lipids are produced in and extruded from lamellar granules as described above or are synthesized in the sebaceous glands and then excreted to the skin’s surface through the hair follicle. The excretion of sebum by sebaceous glands is hormonally controlled (see Chapter 10). Lipids help keep the NMF inside the cells where it is needed to keep cells hydrated and aqueous enzymes functioning. Although this is less well characterized, lipids can themselves influence enzyme function.
ROLE OF LIPIDS IN TEWL
1-5 The keratinocytes are embedded in a lipid matrix that resembles bricks and mortar. Natural moisturizing factor (NMF) is present within the keratinocytes.NMF and the lipid bilayer prevent dehydration of the epidermis.
The major lipids found in the SC that contribute to the water permeability barrier are ceramides, cholesterol, and fatty acids. Since the 1940s, when the SC was first identified as the primary barrier to water loss, many hypotheses have been entertained as to exactly which lipids are important in the SC. The research with the EFAD mice described above led to a focus on phospholipids because they contain linoleic acid. However, it was later found that phospholipids are almost completely absent from the SC.40 In 1982, ceramide 1 was discovered. This lipid compound is rich in linoleic acid and is believed to play a major role in structuring SC lipids essential for barrier function.45 Later, five more distinct types of ceramides were discovered and named according to the polarity of the molecule. Ceramide 1 is the most nonpolar and ceramide 6 is the most polar. Although the ceramides were once thought to be the key to skin moisturization, studies now suggest that no particular lipid is more important than the others. It appears that the proportion of fatty acids, ceramides, and cholesterol is the most important parameter. This was demonstrated in a study in which after altering the water barrier with acetone, the application of a combination of ceramides, fatty acids, and cholesterol resulted in normal barrier recovery.46 Application of each of the separate entities alone resulted in delayed barrier recovery. Manufacturers now include ceramides or a mixture of ceramides, cholesterol, and fatty acids in several available products as a result of these findings. However, the use of these mixtures to
FIGURE
DEEP
SUPERFICIAL
Corneocytes (bricks)
treat atopic dermatitis and other ichthyotic disorders has been disappointing.
SUMMARY
The epidermis is implicated in many of the skin complaints of cosmetic patients. It is the state of the epidermis that causes the skin to feel rough and appear dull. A flexible, well-hydrated epidermis is more supple and radiant than a dehydrated epidermis. The popularity of buff puffs, exfoliating scrubs, masks, moisturizers, chemical peels, and microdermabrasion attest to the obsession that cosmetic patients have with the condition of their epidermis. It is important to understand the properties of the epidermis in order to understand which cosmetic products and procedures can truly benefit patients as opposed to those that are based on myths or hype.
REFERENCES
1.Chu D. Overview of biology, development, and structure of skin. In: Wolff K, Goldsmith L, Katz S, Gilchest B, Paller A, Leffell D, eds. Fitzpatrick’s Dermatology in General Medicine. 7th ed. New York, NY: Mcgraw-Hill; 2008:60.
2.Braff MH, Di Nardo A, Gallo RL. Keratinocytes store the antimicrobial peptide cathelicidin in lamellar bodies.
J Invest Dermatol. 2005;124:394.
3.Li L, Tucker RW, Hennings H, et al. Inhibitors of the intracellular Ca(2+)ATPase in cultured mouse keratinocytes reveal components of terminal differentiation that are regulated by distinct intracellular Ca2+ compartments. Cell Growth Differ. 1995;6:1171.
4.Green H. The keratinocyte as differentiated cell type. Harvey Lect. 1980;74:101.
5.Eckert RL, Crish JF, Robinson NA. The epidermal keratinocyte as a model for the study of gene regulation and cell differentiation. Physiol Rev. 1997;77:397-424.
6.Su MJ, Bikle DD, Mancianti ML, et al. 1,25-Dihydroxyvitamin D3 potentiates the keratinocyte response to calcium.
J Biol Chem. 1994;269:14723.
7.Hosomi J, Hosoi J, Abe E, et al. Regulation of terminal differentiation of cultured mouse epidermal cells by 1 alpha, 25-dihydroxyvitamin D3 Endocrinology. 1983;113:1950.
8.Smith EL, Walworth NC, Holick MF. Effect of 1 alpha,25-dihydroxyvitamin D3 on the morphologic and biochemical differentiation of cultured human epidermal keratinocytes grown in serum-free conditions. J Invest Dermatol. 1986;86:709.
9.Yuspa SH, Kilkenny AE, Steinert PM, et al. Expression of murine epidermal differentiation markers is tightly regulated by restricted extracellular calcium concentrations in vitro. J Cell Biol. 1989;109:1207.
10.Sharpe GR, Gillespie JI, Greenwell JR. An increase in intracellular free calcium is an early event during differentiation of cul-
tured human keratinocytes. FEBS Lett 1989;254:25.
11.Oda Y, Sihlbom C, Chalkley RJ, et al. Two distinct coactivators, DRIP/mediator and SRC/p160, are differentially involved in VDR transactivation during keratinocyte differentiation. J Steroid Biochem Mol Biol. 2004;273:89-90.
12.Bikle D, Teichert A, Hawker N, et al. Sequential regulation of keratinocyte differentiation by 1,25(OH)2D3, VDR, and its coregulators. J Steroid Biochem Mol Biol. 2007;103:396.
13.Christophers E, Kligman AM. Visualization of the cell layers of the stratum corneum. J Invest Dermatol. 1964;42:407.
14.Blair C. Morphology and thickness of the human stratum corneum. Br J Dermatol 1968;80:430.
15.Proksch E, Jensen J. Skin as an organ of protection. In: Wolff K, Goldsmith L, Katz S, Gilchest B, Paller A, Leffell D, eds. Fitzpatrick’s Dermatology in General Medicine . 7th ed. New York, NY: McGraw-Hill; 2008:383-395.
16.Egelrud T. Desquamation. In: Loden M, Maibach H, eds. Dry Skin and Moisturizers 1st ed. Boca Raton, FL: CRC Press; 2000:110.
17.Proksch E, Jensen J. Skin as an organ of protection. In: Wolff K, Goldsmith L, Katz S, Gilchest B, Paller A, Leffell D, eds. Fitzpatrick’s Dermatology in General Medicine . 7th ed. New York, NY: McGraw-Hill; 2008:87.
18.Yaar M, Gilchrest B. Aging of skin. In: Freedberg IM, Eisen A, Wolff K, Austen K, Goldmsith L, Katz S, Fitzpatrick T, eds. Fitzpatrick’s Dermatology in General Medicine . 5th ed. New York, NY: McGraw-Hill; 1999:1697-1706.
19.Jost M, Kari C, Rodeck U. The EGF receptor—an essential regulator of multiple epidermal functions. Eur J Dermatol 2000;10:505.
20.Miki T, Bottaro DP, Fleming TP, et al. Determination of ligand-binding specificity by alternative splicing: two distinct growth factor receptors encoded by a single gene. Proc Natl Acad Sci U.S.A 1992;89:246.
21.Brauchle M, Fässler R, Werner S. Suppression of keratinocyte growth factor expression by glucocorticoids in vitro and during wound healing. J Invest Dermatol. 1995;105:579.
22.Karvinen S, Pasonen-Seppänen S, Hyttinen JM, et al. Keratinocyte growth factor stimulates migration and hyaluronan synthesis in the epidermis by activation of keratinocyte hyaluronan synthases 2 and 3. J Biol Chem. 2003;278:49495.
23.William I, Rich B, Kupper T. Cytokines. In: Wolff K, Goldsmith L, Katz S, Gilchest B, Paller A, Leffell D, eds. Fitzpatrick’s Dermatology inGeneral Medicine. 7th ed. New York, NY: McGraw-Hill; 2008:116.
24.Shah M, Foreman DM, Ferguson MW. Neutralisation of TGF-beta 1 and TGFbeta 2 or exogenous addition of TGFbeta 3 to cutaneous rat wounds reduces scarring. J Cell Sci. 1995;108:985.
25.Shah M, Foreman DM, Ferguson MW. Control of scarring in adult wounds by neutralising antibody to transforming growth factor beta. Lancet . 1992; 339:213.
27.Izadpanah A, Gallo RL. Antimicrobial peptides. J Am Acad Dermatol. 2005; 52:381.
28.Frohm M, Agerberth B, Ahangari G, et al. The expression of the gene coding for the antibacterial peptide LL-37 is induced in human keratinocytes during inflammatory disorders. J Biol Chem. 1997;272: 15258.
29.Murakami M, Ohtake T, Dorschner RA, et al. Cathelicidin anti-microbial peptide expression in sweat, an innate defense system for the skin. J Invest Dermatol 2002;119:1090.
30.Murakami M, Ohtake T, Dorschner RA, et al. Cathelicidin antimicrobial peptides are expressed in salivary glands and saliva. J Dent Res.2002;81:845.
31.Heilborn JD, Nilsson MF, Kratz G, et al. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J Invest Dermatol 2003;120:379.
32.Gallo RL, Ono M, Povsic T, et al. Syndecans, cell surface heparan sulfate proteoglycans, are induced by a prolinerich antimicrobial peptide from wounds. Proc Natl Acad Sci U S A. 1994;91:11035.
33.Ali RS, Falconer A, Ikram M, et al. Expression of the peptide antibiotics human beta defensin-1 and human beta defensin-2 in normal human skin. J Invest Dermatol. 2001;117:106.
34.Mathews M, Jia HP, Guthmiller JM, et al. Production of beta-defensin antimicrobial peptides by the oral mucosa and salivary glands. Infect Immun. 1999;67:2740.
35.Dunsche A, Acil Y, Dommisch H, et al. The novel human beta-defensin-3 is widely expressed in oral tissues. Eur J Oral Sci. 2002;1110:121.
36.Frye M, Bargon J, Gropp R. Expression of human beta-defensin-1 promotes differentiation of keratinocytes. J Mol Med 2001;79:275.
37.Seo SJ, Ahn SW, Hong CK, et al. Expressions of beta-defensins in human keratinocyte cell lines. J Dermatol Sci. 2001;27:183.
38.Sybert VP, Dale BA, Holbrook KA. Ichthyosis vulgaris: identification of a defect in synthesis of filaggrin correlated with an absence of keratohyaline granules. J Invest Dermatol. 1985;84:191.
39.Scott IR, Harding CR. Physiological effects of occlusion-filaggrin retention (abstr). Dermatology. 1993;2000:773.
40.Rawlings AV, Scott IR, Harding CR, et al. Stratum corneum moisturization at the molecular level. J Invest Dermatol. 1994; 103:731.
41.Downing DT, Strauss JS, Pochi PE. Variability in the chemical composition of human skin surface lipids. J Invest Dermatol. 1969;53:322.
42.Webster D, France JT, Shapiro LJ, et al. Xlinked ichthyosis due to steroid-sulphatase deficiency. Lancet. 1978;1:70.
43.Prottey C. Essential fatty acids and the skin. Br J Dermatol. 1976;94:579.
45.Swartzendruber DC, Wertz PW, Kitko DJ, et al. Molecular models of the intercellular lipid lamellae in mammalian stratum corneum. J Invest Dermatol. 1989;92:251.
46.Man MQ, Feingold KR, Elias PM. Exogenous lipids influence permeability barrier recovery in acetone-treated murine skin. Arch Dermatol. 1993;129:728.
CHAPTER 2
Basic Science of the Dermis
Leslie Baumann,MD
Sogol Saghari,MD
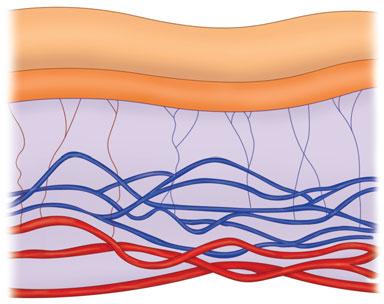
The dermis lies between the epidermis and the subcutaneous fat. It is responsible for the thickness of the skin, and as a result plays a key role in the cosmetic appearance of the skin. The thickness of the dermis varies over different parts of the body and the size doubles between the ages of 3 and 7 years and again at puberty. With aging, this basic layer decreases in thickness and moisture. The dermis, which is laden with nerves, blood vessels, and sweat glands, consists mostly of collagen. The uppermost portion of this layer, which lies beneath the epidermis, is known as the papillary dermis and the lower portion is known as the reticular dermis. Smaller collagen bundles, greater cellularity, and a higher density in its vascular elements characterize the papillary dermis as compared to the reticulardermis. Fibroblasts are the primary cell type in the dermis. They produce collagen, elastin, other matrix proteins, and enzymes such as collagenase and stromelysin. These structural components will be discussed individually because each exhibits significant characteristics that influence the function of the skin. Immune cells such as mast cells, polymorphonuclear leukocytes, lymphocytes, and macrophages are also present in the dermis.
The junction between the epidermis and dermis is known as the dermal–epidermal junction (DEJ) (Fig. 2-1). Much
is known about the attachment proteins found in the basement membrane of the DEJ. At this point there are no known cosmetic implications for this area, as such a discussion is beyond the scope of this book. Instead, this chapter will focus on the components of the dermis that are known to be important in aging.
COLLAGEN
Collagen, one of the strongest natural proteins and the most abundant one in humans as well as in skin, imparts durability and resilience to the skin. It has been the focus of much antiaging research and the target of several skin products and procedures. The importance of collagen is emphasized in the literature regarding many of the topical agents that are touted to increase collagen synthesis such as glycolic and ascorbic acids. Resurfacing techniques such as the CO2 laser and dermabrasion are intended to change collagen structure, thereby improving skin texture. Various forms of collagen are injected into the dermis to replace damaged collagen and to reverse the signs of aging. Finally, topical retinoids have been shown to reduce the collagen damage that occurs because of sun exposure. These sundry
aspects of collagen health or replacement will be discussed separately in upcoming chapters; however, it is necessary first to gain an understanding of the structure and function of collagen.
“Collagen” is actually a complex family of 18 proteins, 11 of which are present in the dermis. Collagen fibers are always seen in the dermis in the final, mature state of assembly as opposed to elastin, the immature fibers of which are seen in the superficial dermis with the more mature fibers found in the deeper layer of the dermis. Each type of collagen is composed of three chains (Fig. 2-2) . Collagen is synthesized in the fibroblasts in a precursor form called procollagen. Proline residues on the procollagen chain are converted to hydroxyproline by the enzyme prolyl hydroxylase. This reaction requires the presence of Fe++, ascorbic acid (vitamin C), and -ketoglutarate. Lysine residues on the procollagen chain are also converted to hydroxylysine; in this case, by the enzyme lysyl hydroxylase. This reaction also requires the presence of Fe ++ , ascorbic acid, and -ketoglutarate. It is interesting to note that a deficiency of vitamin C, which is an essential mediating component in these reactions, leads to scurvy, a disease characterized by decreased collagen production.
Collagen Glycation
Glycation of extracellular matrix (ECM) collagen and proteins plays an important role in the aging process. This is not to be confused with glycosylation of collagen, which is an enzyme-mediated process in the intracellular step of collagen biosynthesis. Glycation is a nonenzymatic series of biologic events that involves adding a reducing sugar molecule (such as glucose or fructose) to ECM collagen and proteins. This reaction is also known as the Maillard reaction. The sugar molecule mainly reacts with the amino group side chains
FIGURE 2-1 Histopathology of the dermal-epidermal junction.The basement membrane separates the epidermis and the dermis.(Image courtesy of George Loannides,MD.)
Basement membrane
FIGURE 2-2 Collagen is formed when three chains come together to form a triple helix.
Glycation End Products (AGEs)
of lysine and arginine of collagen and ECM proteins. Subsequently, the product of this process undergoes oxidative reactions resulting in the formation of advanced glycation end products (AGEs) (Fig. 2-3) . AGEs have been implicated in the aging process and agerelated diseases such as diabetes mellitus, 1–3 chronic renal failure, 4,5 and Alzheimer’s disease. 6–8 It is believed that with time, AGEs increase,9 accumulate on human collagen10 and elastin fibers,11 and contribute to aging of the skin. As a result of glycation, collagen networks lose their ability to contract, and they become stiffer and resistant to remodeling. Fibroblasts are key elements for collagen contracture, as they apply contracture force on the collagen lattice via their actin cytoskeleton. 12 Glycated collagen modifies the actin cytoskeleton of the fibroblasts thereby diminishing their collagen contraction capacity.13 Fibroblasts also secrete collagenase (MMP-1), which is essential for collagen turnover. Glycated collagen has been proven to decrease levels of collagenase I (MMP-1), leading to less tissue remodeling.14 Studies have shown that UV exposure may also contribute to the production and function of AGEs. N e -(carboxymethyl) lysine
Major Collagen Types Found in the Dermis
(CML) is one of the AGEs in which the amino side chain of lysine is reduced. This product was shown to accumulate on elastin tissue of photoaged skin and proven to be higher in sun-exposed skin as compared to sun-protected skin.11 In addition, it has been proposed that AGE-modified proteins act as endogenous photosensitizers in human skin via oxidative stress mechanisms induced by UVA light.15
The Key Types of Collagen Found in the Dermis (Table 2-1)
Type I collagen comprises 80% to 85% of the dermal matrix and is responsible for the tensile strength of the dermis. The amount of collagen I has been shown to be lower in photoaged skin, and to be increased after dermabrasion procedures.16 Therefore, it is likely that collagen I is the most important collagen type in regard to skin aging. Type III is the second most important form of collagen in the dermis, making up anywhere from 10% to 15% of the matrix. 17 This collagen type has a smaller diameter than type I and forms smaller bundles allowing for skin pliability. Type III, also known as “fetal collagen” because it predominates in
embryonic life, is seen in higher amounts around the blood vessels and beneath the epidermis.
The other types of collagen that are noteworthy for a cosmetic dermatologist are type IV collagen, which forms a structure lattice that is found in the basement membrane zone and type V collagen, which is diffusely distributed throughout the dermis and comprises roughly 4% to 5% of the matrix. Type VII collagen makes up the anchoring fibrils in the DEJ. Type XVII collagen is located in the hemidesmosome and plays an important structural role as well. The importance of these collagens and other structural proteins is evident in genetic diseases characterized by a lack of these structures and in acquired diseases characterized by antibody formation to these important structures. For example, patients with an inherited blistering disease known as dominant dystrophic epidermolysis have been shown to have a scarcity of type VII collagen with resulting abnormalities in their anchoring fibrils. An acquired bullous disease, epidermolysis bullosa acquisita (EBA), is caused by antibodies to this same collagen type VII. Although the discussion of these diseases is beyond the scope of this text, it is interesting that patients with chronic sun exposure have also been found to have alterations in collagen type VII. This may contribute to the skin fragility seen in elderly patients. Some investigators have postulated that a weakened bond between the dermis and epidermis caused by loss of the anchoring fibrils (collagen VII) may lead to wrinkle formation.18 The importance of collagen and changes seen in aged skin will be discussed further in Chapter 6.
ELASTIN
Elastic fibers represent one of the essential components of the ECM of connective tissue (Fig. 2-4). They confer resilience
I Bone,tendon,skin Gives tensile strength80
III Fetal collagenDermis,GI,vessels Gives compliance15
IV Basement membranesForms a lattice
V Dermis,diffusely distributedUnknown 4–5 epidermolysis bullosa
VII Anchoring fibrils Stabilizes DEJ acquisita (EBA),dystrophic
FIGURE 2-3 Glycation of proteins is thought to play a role in the aging process.
TABLE 2-1
and elasticity to skin as well as other organs such as the lungs and blood vessels. Elastogenesis starts during fetal life and reaches its maximum near birth and the early neonatal period. It then decreases significantly and is virtually nonexistent by adult life. Elastic fibers have two components. Their main component is elastin, an amorphous, insoluble connective tissue protein. Elastin is surrounded by microfibrils, the second component. Elastin constitutes 2% to 3% of the dry weight of skin, 3% to 7% of lung, 28% to 32% of major blood vessels, and 50% of elastic ligaments.19
Elastin is produced from its precursor tropoelastin in the fibroblasts as well as endothelial cells and vascular smooth muscle cells. In contrast to collagen fibers, elastin fibers are present in the dermis in various levels of maturity. The least mature fibers are called oxytalan. They course perpendicularly from the DEJ to the top of the reticular dermis. More mature elastin fibers, called elaunin, then attach to a horizontal plexus of fibers found in the reticular dermis. Elaunin is more mature because it has more elastin deposited on the fibrillin mesh. The most mature elastin fibers are unnamed and are found deeper in the reticular dermis (Fig. 2-5)
Microfibrils play a very important role in elastogenesis and act as a scaffold for tropoelastin deposition and assembly.20 Microfibrils are primarily composed of glycoproteins from the fibrillin family and microfibril-associated glycoprotein (MAGP)-1 and -2. Fibrillin-1 has been shown to be important in elastic fiber development21 and wound repair.22 Microfibrils are adjacent to tropoelastinproducing cells and parallel to the developing elastin fiber.23 The microfibrils form a template on which tropoelastin is deposited. The tropoelastin polypeptides are then covalently cross-linked to form elastin. Tropoelastin polypeptides contain alternating hydrophilic and hydrophobic regions. The hydrophobic
domains, which are rich in proline, valine, and glycine, are believed to be responsible for the elasticity of the elastin tissue.24 The hydrophilic domains on the other hand are rich in alanine and lysine, and interact with the enzyme lysyl oxidase in the process of cross-linking.25 The cross-linking of elastin is a complex process necessary for its proper function and stability. This process is mediated via the copperrequiring enzyme lysyl oxidase,26 and the subsequent formation of desmosine and isodesmosine cross-links, which result in an insoluble elastin network.27 Elastin is fascinating and although much is known about it, its relevance in cosmetic dermatology is unclear. It seems certain that collagen, hyaluronic acid (HA), and elastin bind each other covalently and make up a three-dimensional structure that is impaired in aged skin. There is a commonly held belief that these three components must be increased in order to give skin a younger appearance. However, the trick is that de novo elastin production does not occur in adulthood. Trying to increase production of elastin in adults will surely be a focus of cosmetic dermatology research in the future.
The elastic fiber’s structure provides clues about its ability to interact with HA and collagen. Mature elastic fibers contain an array of proteoglycans. Versican is one of the most widely studied proteoglycans28 and is a member of the hyaluronan binding family that also includes aggrecan and neurocan. Versican contributes to cell adhesion, proliferation, and migration and can interact with multiple ECM proteins to mediate assembly. Mature elastic fibers are found at the periphery of collagen bundles, offering a clue that elastin has important interactions with collagen as well as with HA.
Elastic fibers are degraded by the elastolytic enzymes such as human leukocyte elastase (HLE). With significant levels of sun exposure, elastin degrades and is seen as an amorphous substance in the dermis when viewed by light microscopy. This resultant “elastosis” is a hallmark of photoaged skin. Interestingly, there are protective mechanisms in the skin preventing elastin degradation. Lysozymes are believed to play a protective role in this matter. They have been shown to increase and deposit on the elastin fibers of UVexposed skin.29 By binding to the elastin, the lysozymes prevent the proper interaction between elastase and elastin,30 thereby inhibiting the proteolytic activity of the elastolytic enzymes.30,31 It is also believed that damage to the elastin fibers leads to the decreased skin elasticity seen in aged skin.32 Defects or damage to elastin may lead to wrinkles even in the absence of sun exposure and aging. Indeed, in one case, a child with “wrinkled skin syndrome” was shown to have a deficiency of elastin fibers,33 which demonstrates the importance of elastin in skin integrity. Defective elastic fibers can give rise to multiple dermatologic diseases including cutis laxa, pseudoxanthoma elasticum
illary dermis
fibers
icular dermis
in fibers
reticular dermis
FIGURE 2-5 The elastic fiber network in the dermis consists of immature oxytalan fibers in the superficial dermis and the more mature elaunin fibers in the middle dermis.The most mature elastic fibers are unnamed and are found in the deep reticular dermis.
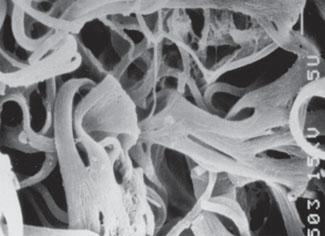
FIGURE 2-4 A and B.Scanning electron micrographs of the elastic fibers in human skin.Adapted from Fitzpatrick’s Dermatology in General Medicine,seventh edition (McGraw Hill),page 532,with permission.
EPIDERMIS DE Junction
(PXE), elastosis perforans serpiginosa (also known as Lutz-Miescher’s syndrome), and dermatofibrosis lenticularis (also known as Buschke-Ollendorf syndrome).
Studies have demonstrated a reduction in the elastin content in protected areas of the skin with aging. In a study performed on Egyptian subjects, the relative amount of elastin in the non-UVexposed abdominal skin significantly decreased from 49.2% 0.6% in the first decade to 30.4% 0.8% in the ninth decade.34 Another study on elastin content in the nonexposed buttock skin of 91 Caucasians between 20 and 80 years of age showed a reduction of 51% in elastin tissue.31 Although UV exposure may result in elastosis and a higher content of elastin tissue, the elastic fibers are rendered structurally abnormal,34 which is microscopically seen as thickened and twisted granular deposits of elastin in the dermis.
Replacing the elastin component of the ECM has always posed a challenge in skin rejuvenation approaches. Researchers have investigated the production of recombinant and cross-linked tropoelastin in great detail.35 However, since it is very difficult to have elastin pass through human skin, stimulating the dermis to produce elastin may be an alternative option. Recently, zinc has become a subject of interest as an elastin tissue stimulator in the skin. Zinc has been shown to increase the epidermal growth factor (EGF) receptor signaling pathway.36 It increases protein tyrosine phosphorylation by inhibiting protein tyrosine phosphatase (PTPase),37 and activates mitogen-activated protein (MAP) kinases,38 which are important for cosignaling in ECM production. Clinical studies have suggested improvement in the elasticity of periocular skin following use of a patented zinc complex topical preparation.39 In a 4-week study of 27 female subjects with a zinc complex-containing eye product, overall improvement of the eye area was noted by 78%, reduction of fine lines by 74%, and firmer skin by 70% of the patients.39 These studies, although promising, need to be conducted in a larger patient population.
GLYCOPROTEINS
Glycoproteins (GP) influence cell migration, adhesion, and orientation. Fibronectin and tenascin are the GPs most relevant in the dermis although vitronectin, thrombospondin, and epi-
bolin are also present in the dermis. Fibronectin is a filamentous GP that mediates platelet binding to collagen, development of granulation tissue, and reepithelialization. Chemotactic for monocytes, fibronectin contains six binding sites including one for collagen, two for heparin, and a region that binds fibrin. Tenascin is abundant in developing skin but found only in the papillary dermis in adult skin. These matrix components play a significant role in tissue remodeling and are important in wound healing following cosmetic procedures.
GLYCOSAMINOGLYCANS
Glycosaminoglycans (GAGs) are polysaccharide chains composed of repeating disaccharide units linked to a core protein. Together the GAGs and attached core protein form proteoglycans. All GAGs except for HA are synthesized in Golgi apparatus. HA is the only GAG that is not produced on a core protein; rather, it is synthesized by an enzyme complex of the plasma membrane.40
Although all the functions of GAGs are not understood, it is known that these compounds avidly bind water and may contribute to the maintenance of salt and water balance. GAGs are found in areas with a fibrous matrix where cells are closely associated but have little space for free movement. Most studies on human skin show an age-related decline in GAG content. The most abundant GAGs in the dermis are HA, which is the only nonsulfated GAG, and dermatan sulfate. The other GAGs include heparin sulfate, heparin, keratan sulfate, chondroitin-4, and chondroitin-6-sulfate. HA is a very important component of the dermis that is responsible for attracting water and giving the dermis its volume. The name reflects its glassy appearance (the Greek word for glass is hyalos) and the presence of a sugar known as uronic acid. HA is known to be important in cell growth, membrane receptor
function, and adhesion. Its structure is identical, whether it is derived from bacterial cultures, animals, or humans (Fig. 2-6). HA appears freely in the dermis and is more concentrated in areas where cells are less densely packed. In young skin, HA is found at the periphery of collagen and elastin fibers and at the interface of these types of fibers. These connections with HA are absent in aged skin.41 HA is a popular ingredient in cosmetic products because it acts as a humectant. Several types are also available in an injectable version for the treatment of wrinkles (see Chapter 23). HA appears to also play a role in keratinocyte differentiation and formation of lamellar bodies via its interaction with CD44,42 a cell surface glycoprotein receptor with HA binding sites.43–45
Decorin is a member of the small leucine-rich proteoglycans (SLRPs) found in the extracelluar matrix protein. Its name is derived from its apparent “decorating” of collagen fibers. Decorin contains a core protein with a high content of leucine repeats and GAG chains of dermatan or chondroitin sulfate. It is shaped in a “horseshoe” pattern and binds to collagen fibrils, resulting in their proper organization.46
Decorin-deficient mice have shown clinical skin fragility and irregular collagen fibrils with increased interfibrillar space on histology.47 In addition to collagen fibrillogenesis, decorin interacts with fibronectin48 and fibrinogen,49 thereby playing a role in wound healing and hemostasis. Another interesting function of decorin is that it reduces the proliferation of cells in neoplasms by stopping their growth in the G1 phase of the cell cycle.50 Carrinoet al.51 studied the catabolic fragment of decorin in adult skin. They noted a higher content of the altered decorin in adult dermis as opposed to nonmeasurable amounts in fetal skin and named it “decorunt.” Decorunt was shown to have a lower affinity for collagen fibrils. This finding may explain some of the changes related to collagen disorganization in aging skin.
FIGURE 2-6 HA is made of repeating dimers of glucuronic acid and N-acetyl glucosamine assembled into long chains.
MATRIX METALLOPROTEINASES
The ECM architecture of human skin is based on its continuous remodeling. This process requires ECM-degrading enzymes followed by synthesis and deposition of new molecules. The matrix metalloproteinases (MMPs), which include a large family of zincdependent endopeptidases, are crucial to the turnover of ECM components. Interstitial collagenase, or MMP-1, was the first enzyme discovered in this group. MMP-1 is secreted from the fibroblasts and is mainly involved in the degradation of collagen types I, II, and III, but has been shown to also cleave the anchoring fibrils of collagen VII.52
Human neutrophil collagenase (MMP8), another type of collagenase, is engaged in cleaving collagen types I and III. Collagenase 3 (MMP-13) is the third member of this group of enzymes, and it is known to fragment fibrillar collagens. It is also believed to have a role in scarless wound healing53 by enhancing fibroblast proliferation and survival.54 Gelatinases are another class of MMPs and consist of two types of enzymes, gelatinase A (MMP-2) and gelatinase B (MMP-9), that are responsible for attacking gelatin and collagen IV in the basement membrane. Other groups of MMPs include stromelysins, which are mainly involved in degradation of proteoglycans, laminins, collagen IV, and matrilysin, which is expressed on stromal tissue, fetal skin, and in the setting of carcinomas.55
The activity of MMPs is regulated by an endogenous tissue inhibitor of metalloproteinases (TIMPs). TIMPs are naturally produced proteins that specifically inhibit the MMPs. The balance between MMPs and their inhibition by TIMPs leads to proper tissue remodeling. TIMPs are regulated via expression of cytokines (such as IL-1), growth factors, and even retinoids.56,57 Retinoids have been shown to provoke a two- to threefold increase in the biosynthesis of human fibroblast-derived TIMP in vitro.58 Increased production of MMPs and decreased production of TIMPs have a role in the metastatic behavior of tumors. Synthetic inhibitors of MMPs are of interest to researchers especially in the area of cancer research. These inhibitors, such as hydroxamates, contain a zinc-chelating group that binds to the active site of MMPs leading to its inhibition. Currently, their use is mostly limited to research studies because of their side-effect profile. Certain medications such as doxycycline are also
known for their inhibitory effect on MMPs and have been studied in myriad MMP-related conditions such as periodontal and atherosclerotic diseases.
HYPODERMIS
The hypodermis, or subcutis, located beneath the dermis, is composed mostly of fat, which is an important energy source for the body. This layer also contains collagen types I, III, and V. As humans age, some of the subcutaneous fat is lost or redistributed into undesired areas. This phenomenon contributes to the aged appearance. Fat injections have been employed to move fat from undesired areas into desired areas where fat has been lost, such as the lower face (see Chapter 23).
The adipocytes secrete a hormone called leptin, a product of the obesity (ob) gene. Leptin exhibits a regulatory effect on human metabolism and appetite and therefore affects adipose tissue mass. Leptin has been shown to be higher in the serum of obese patients, with commensurate levels found in body fat percentage.59 It is believed that a higher percentage of body fat results in elevated leptin levels and the turning off of signals to the brain for appetite reduction. Recombinant leptin injections in mice have been associated with reduction of weight and body fat percentage.60 However, more research is needed to ascertain the therapeutic potential of leptin in humans.
SUMMARY
Although the epidermis is the target of most topical cosmetic products because most do not penetrate to the dermis, the dermis is the target for many of the injectable treatments for aging. The dermis is an extremely important component in skin appearance because it is responsible for imparting thickness and suppleness to the skin. A thinner dermis and an altered DEJ are hallmarks of aged skin. Loss of collagen, elastin, and GAGs located primarily in the dermis contribute significantly to cutaneous aging. Various measures intended to prevent or retard aging target these key constituents of the dermis.
REFERENCES
1.Monnier VM, Kohn RR, Cerami A. Accelerated age-related browning of human collagen in diabetes mellitus. Proc Natl Acad Sci U S A. 1984;81:583.
2.Schnider SL, Kohn RR. Glucosylation of human collagen in aging and diabetes mellitus. J Clin Invest. 1980;66:1179.
3.Schnider SL, Kohn RR. Effects of age and diabetes mellitus on the solubility and nonenzymatic glucosylation of human skin collagen. J Clin Invest. 1981;67:1630.
4.Yamada K, Miyahara Y, Hamaguchi K, et al. Immunohistochemical study of human advanced glycosylation endproducts (AGE) in chronic renal failure. Clin Nephrol. 1994;42:354.
5.Thornalley PJ. Advanced glycation end products in renal failure. J Ren Nutr 2006;16:178.
6.Vitek MP, Bhattacharya K, Glendening JM, et al. Advanced glycation end products contribute to amyloidosis in Alzheimer disease. Proc Natl Acad Sci U S A. 1994;91:4766.
7.Yan SD, Chen X, Schmidt AM, et al. Glycated tau protein in Alzheimer disease: a mechanism for induction of oxidant stress. Proc Natl Acad Sci U S A 1994;91:7787.
8.Takeuchi M, Kikuchi S, Sasaki N, et al. Involvement of advanced glycation endproducts (AGEs) in Alzheimer’s disease. Curr Alzheimer Res. 2004;1:39.
9.Dyer DG, Dunn JA, Thorpe SR, et al. Accumulation of Maillard reaction products in skin collagen in diabetes and aging. J Clin Invest. 1993;91:2463.
10.Verzijl N, DeGroot J, Odehinkel E, et al. Age-related accumulation of Maillard reaction products in human articular cartilage collagen. Biochem J. 2000;350: 381.
11.Mizutari K, Ono T, Ikeda K, et al. Photoenhanced modification of human skin elastin in actinic elastosis by N(epsilon)(carboxymethyl)lysine, one of the glycoxidation products of the Maillard reaction. J Invest Dermatol. 1997;108:797.
12.Tomasek JJ, Haaksma CJ, Eddy RJ, et al. Fibroblast contraction occurs on release of tension in attached collagen lattices: dependency on an organized actin cytoskeleton and serum. Anat Rec. 1992; 232:359.
13.Howard EW, Benton R, Ahern-Moore J, et al. Cellular contraction of collagen lattices is inhibited by nonenzymatic glycation. Exp Cell Res. 1996;228:132.
14.Rittie L, Berton A, Monboisse JC, et al. Decreased contraction of glycated collagen lattices coincides with impaired matrix metalloproteinase production. Biochem Biophys Res Commun. 1999;264:488.
15.Wondrak GT, Roberts MJ, Jacobson MK, et al. Photosensitized growth inhibition of cultured human skin cells: mechanism and suppression of oxidativestress from solar irradiation of glycated proteins. J Invest Dermatol. 2002;119:489.
16.Nelson B, Majmudar G, Griffiths C, et al. Clinical improvement following dermabrasion of photoaged skin correlates with synthesis of collagen I. Arch Derm. 1994;130:1136.
17.Oikarinen A. The aging of skin: chronoaging versus photoaging. Photodermatol Photomed. 1990;7:3.
18.Craven NM, Watson RE, Jones CJ, et al. Clinical features of photodamaged human skin are associated with a reduction in collagen VII. Br J Derm. 1997;137: 344.
20.Robb BW, Wachi H, Schaub T, et al. Characterization of an in vitro model of elastic fiber assembly. Mol Biol Cell. 1999; 10:3595.
21.Kielty CM, Sherratt MJ, Shuttleworth CA. Elastic fibres. J Cell Sci. 2002;115: 2817.
22.Amadeu TP, Braune AS, Porto LC, et al. Fibrillin-1 and elastin are differentially expressed in hypertrophic scars and keloids. Wound Repair Regen. 2004;12: 169.
23.Mithieux SM, Weiss AS. Elastin. Adv Protein Chem. 2005;70:437.
24.Li B, Daggett V. Molecular basis for the extensibility of elastin. J Muscle Res Cell Motil. 2002;23:561.
25.Rosenbloom J, Abrams WR, Mecham R. Extracellular matrix 4: the elastic fiber. FASEB J. 1993;7:1208.
26.Smith-Mungo LI, Kagan HM. Lysyl oxidase: properties, regulation and multiple functions in biology. Matrix Biol 1998;16:387.
27.Starcher BC. Determination of the elastin content of tissues by measuring desmosine and isodesmosine. Anal Biochem 1977;79:11.
28.Wight TN. Versican: a versatile extracellular matrix proteoglycan in cell biology. Curr Opin Cell Biol. 2002;14:617.
29.Suwabe HA, Serizawa H, Kajiwara M, et al. Degenerative processes of elastic fibers in sun-protected and sun-exposed skin: immunoelectron microscopic observation of elastin, fibrillin-1, amyloid P component, lysozyme and alpha1antitrypsin. Pathol Int. 1999;49: 391.
30.Park PW, Biedermann K, Mecham L, et al. Lysozyme binds to elastin and protects elastin from elastase-mediated degradation. J Invest Dermatol. 1996;106:1075.
31.Seite S, Zucchi H, Septier D, et al. Elastin changes during chronological and photoageing: the important role of lysozyme. J Eur Acad Dermatol Venereol. 2006;20:980.
32.Escoffier C, de Rigal J, Rochefort A, et al. Age-related mechanical properties of human skin: an in vivo study. J Invest Dermatol. 1989;93:353.
33.Boente MC, Winik BC, Asial RA. Wrinkly skin syndrome: ultrastructural alterations of the elastic fibers. Pediatr Dermatol. 1999;16:113.
34.El-Domyati M, Attia S, Saleh F, et al. Intrinsic aging vs. photoaging: a comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp Dermatol. 2002;11:398.
35.Mithieux SM, Wise SG, Raftery MJ, et al. A model two-component system for studying the architecture of elastin assembly in vitro. J Struct Biol. 2005;149:282.
36.Wu W, Graves LM, Jaspers I, et al. Activation of the EGF receptor signaling pathway in human airway epithelial cells exposed to metals. Am J Physiol . 1999; 277:L924.
37.Samet JM, Silbajoris R, Wu W, et al. Tyrosine phosphatases as targets in metal-induced signaling in human airway epithelial cells. Am J Respir Cell Mol Biol. 1999;21:357.
38.Samet JM, Graves LM, Quay J, et al. Activation of MAPKs in human bronchial epithelial cells exposed to metals. Am J Physiol. 1998;275:L551.
39.Baumann L, Weinkle S. Improving elasticity: the science of aging skin. Cosm Dermatol. 2007;20:168.
40.Uitto J, Chu M, Gallo R, Eisen A. Collagen, elastic fibers, and extracellular matrix of the dermis. In: Wolff K, Goldsmith L, Katz S, Gilchrest B, Paller A, Leffell D, eds. Fitzpatrick’s Dermatology in General Medicine. 7th ed. New York, NY: McGraw-Hill; 2008:539.
41.Ghersetich I, Lotti T, Campanile G, et al. Hyaluronic acid in cutaneous intrinsic aging. Int J Dermatol. 1994;33:119.
42.Bourguignon LY, Ramez M, Gilad E, et al. Hyaluronan-CD44 interaction stimulates keratinocyte differentiation, lamellar body formation/secretion, and permeability barrier homeostasis. J Invest Dermatol. 2006;126:1356.
43.Aruffo A, Stamenkovic I, Melnick M, et al. CD44 is the principal cell surface receptor for hyaluronate. Cell. 1990;61:1303.
44.Culty M, Miyake K, Kincade PW, et al. The hyaluronate receptor is a member of the CD44 (H-CAM) family of cell surface glycoproteins. J Cell Biol. 1990;111:2765.
45.Underhill C. CD44: the hyaluronan receptor. J Cell Sci. 1992;103:293.
46.Scott JE. Proteodermatan and proteokeratan sulfate (decorin, lumican/ fibromodulin) proteins are horseshoe shaped. Implications for their interactions with collagen. Biochemistry. 1996;35: 8795.
47.Danielson KG, Baribault H, Holmes DF, et al. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J Cell Biol. 1997; 136: 729.
48.Schmidt G, Robenek H, Harrach B, et al. Interaction of small dermatan sulfate
proteoglycan from fibroblasts with fibronectin. J Cell Biol. 1987;104:1683.
49.Dugan TA, Yang VW, McQuillan DJ, et al. Decorin binds fibrinogen in a Zn2+dependent interaction. J Biol Chem. 2003; 278:13655.
50.De Luca A, Santra M, Baldi A, et al. Decorin-induced growth suppression is associated with up-regulation of p21, an inhibitor of cyclin-dependent kinases. J Biol Chem. 1996;271:18961.
51.Carrino DA, Onnerfjord P, Sandy JD, et al. Age-related changes in the proteoglycans of human skin. Specific cleavage of decorin to yield a major catabolic fragment in adult skin. J Biol Chem. 2003; 278:17566.
52.Seltzer JL, Eisen AZ, Bauer EA, et al. Cleavage of type VII collagen by interstitial collagenase and type IV collagenase (gelatinase) derived from human skin. J Biol Chem. 1989;264:3822.
53.Ravanti L, Hakkinen L, Larjava H, et al. Transforming growth factor-beta induces collagenase-3 expression by human gingival fibroblasts via p38 mitogen-activated protein kinase. J Biol Chem. 1999; 274:37292.
54.Toriseva MJ, Ala-aho R, Karvinen J, et al. Collagenase-3 (MMP-13) enhances remodeling of three-dimensional collagen and promotes survival of human skin fibroblasts. J Invest Dermatol. 2006; 127:49.
55.Karelina TV, Goldberg GI, Eisen AZ. Matrilysin (PUMP) correlates with dermal invasion during appendageal development and cutaneous neoplasia. J Invest Dermatol. 1994;103:482.
56.Reynolds JJ, Hembry RM, Meikle MC. Connective tissue degradation in health and periodontal disease and the roles of matrix metalloproteinases and their natural inhibitors. Adv Dent Res. 1994; 8:312.
57.Wojtowicz-Praga SM, Dickson RB, Hawkins MJ. Matrix metalloproteinase inhibitors. Invest New Drugs. 1997;15:61.
58.Clark SD, Kobayashi DK, Welgus HG. Regulation of the expression of tissue inhibitor of metalloproteinases and collagenase by retinoids and glucocorticoids in human fibroblasts. J Clin Invest. 1987; 80:1280.
59.Considine RV, Sinha MK, Heiman ML, et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med. 1996; 334:292.
60.Pelleymounter MA, Cullen MJ, Baker MB, et al. Effects of the obese gene product on body weight regulation in ob/ob mice. Science. 1995;269:540.
CHAPTER 3
Fat and the Subcutaneous Layer
Voraphol Vejjabhinanta,MD
Suzan Obagi,MD
Anita Singh,MS
Leslie Baumann,MD
Subcutaneous tissue, or the hypodermis, is one of the largest tissues in the human body. The major components of this layer are adipocytes, fibrous tissue, and blood vessels. It is estimated that this layer represents 9% to 18% of body weight in normal-weight men and 14% to 20% in women of normal weight. 1 Fat mass can increase up to four fold in severe obesity, which may represent 60% to 70% of total body weight. 2 Although gaining fat in the body is undesirable for many, losing fat in the face has cosmetic implications as well. Adipose tissue gain and loss and volume changes contribute to the aged appearance of the face and body. This chapter will review the importance of the subcutaneous tissue and its various functions.
The subcutaneous tissue is usually not given as much attention as the dermis and epidermis because pathology at superficial layers is easier to detect or diagnose by a shave or small punch biopsy. Subcutaneous tissue usually must have an extensive defect before it is
BOX 3-1Functions of the Subcutaneous Tissue
•The largest repository of energy in the body.
•Stores fat-soluble vitamins (A,D,E,K), including their derivatives such as retinoic acids.
•Helps to shape the surface of the body,and form fat pads that act as shock absorbers.
•Helps distribute force or stress to mitigate damage to underlying organs.
•Protects against physical injury from excessive heat,cold,or mechanical factors.
•Fills up spaces between other tissues and helps to keep organs in place.
•Involved in thermoregulation by insulating the body from heat loss.
•Functions as a secretory organ that releases many cytokines.
•Plays a role in regulating androgen and estrogen levels.3
noticed, and in order to biopsy this area, an incision or large punch biopsy (e.g., 6 mm) is required. During histologic tissue processing of biopsy tissue, the triglyceride component, which is the major component of adipocytes, is removed by alcohol and xylol. For this reason, subcutaneous tissue has long been ignored. However, with advances in diagnostic methods and new treatments, much more has been learned about the subcutaneous layer (Box 3-1). It is important for dermatologists and cosmetically oriented physicians to pay close attention to this tissue because it has many roles in cosmetic dermatology and general appearance.
ADIPOCYTES
In the past, adipocytes in adults were considered stable, nondividing cells, like other mature cells. However, recent data reveal that adipocytes in adults have the potential to increase in number or revert back into stem cells. These stem calls can differentiate to other tissue, such as fibroblasts, collagen, elastic fibers, and hematopoietic stromal cells.4 Fat cells are derived from undifferentiated fibroblast-like mesenchymal cells. Under certain conditions, these mesenchymal cells give rise to adipose cells. Adipose tissue is classifiedinto two morphologic types: white and brown adipose tissue. White adipose tissue normally appears yellow because of the accumulation of -carotene, while brown adipose tissue was named by its appearance derived from its rich vascular supply. Mature white adipocytes are called round unilocular fat cells. They have a copious supply of cytoplasm, which contains a single, large lipid droplet that pushes the nucleus to the border of the cell. Brown adipocytes, called polygonal multilocular fat cells, have multiple small lipid droplets. When observed with an electron microscope, brown adipocytes demonstrably contain much more mitochondria and smooth endoplasmic reticulum than white adipocytes. In humans, brown adipocytes play a major role in nonshivering thermogenesis. Brown adipose tissue can be found during the fetal and early neonatal phases, while the majority of adipocytes in adults are white adipocytes. Some scientists have tried to elucidate the mechanism of
how brown adipocytes convert fat to energy, in order to find a way to get rid of body fat by stimulating brown fat to return.5
In the past, it was believed that the number of adipocytes, which develop during the 30th week of gestation, does not increase after birth. However, newer evidence has shown that adipocytes can increase in number and size in certain situations or environments. In general, adipocytes are thought to have two periods of growth. The first period occurs from the embryonic stage to 18 months after birth, and the second period occurs during puberty. Changes in adipose tissue mass are determined by both size and number of adipocytes.6 An increase in size (hypertrophy)7 usually precedes an increase in the number of cells (hyperplasia).8
ANATOMY
Subcutaneous tissue, also known as the superficial fascia, is divided into three layers: apical, mantle, and the deeper layer. The apical layer is located beneath the reticular dermis surrounding sweat glands and hair follicles. It contains blood vessels, lymphatic vessels, and nerves. It is also rich in carotenoids and tends to be yellow in gross appearance. Damage to this layer can lead to hematoma, seroma, paresthesia, and full thickness skin necrosis. The mantle layer is composed of columnar-shaped adipocytes and is absent from eyelids, nail beds, bridge of the nose, and penis. It contributes to the ability to resist trauma by distributing pressure across a large field. The deeper layer is located under the mantle layer and its shape depends on gender, genetics, anatomic area, and diet. Adipocytes in this layer are arranged in lobules between septa as well as between fibrous planes. This layer is suitable for liposuction. Vertical extrusion and/or expansion of this layer can cause cellulite (Fig. 3-1).
Subcutaneous tissue is found throughout the body except for the eyelids, proximal nail fold, penis, scrotum, and the entire auricle of the external ear except the lobule. In particular, subcutaneous tissue is prominent at the temples, cheeks, chin, nose, abdomen, buttocks, and thighs, as well as infraorbital areas and very thick at the palms and soles. Age, gender, and lifestyle choices determine the distribution and density of adipose deposits. For example, in newborns
adipose tissue has a uniform thickness throughout the body, while in adults the tissue tends to disappear from some areas of the body and increase in other areas under the influence of hormones.
Adipose tissue is distributed differently in men and women. Men tend to accumulate fat in an android or upper abdominal body distribution (apple shape). In contrast, women tend to accumulate fat in a gynoid or lower body distribution that predominantly involves the lower abdomen, hips, and thighs (pear shape) (Fig. 3-2)
In the elderly, hyper- or hypoaccumulation of fat occurs in various areas. For example, infraorbital eye bags, buccal fat pad accumulation (chipmunk feature), wattle of the anterior neck, loose skin and fat accumulation in the posterior arm, increase in breast size of males, and an increase in abdomen, buttock and thigh fat are common. Subcutaneousfat can also be lost in the
malar fat pad during the aging process. This can lead to prominent flattening of the cheek/buccal area, sagging of the skin of the face, and prominent deep wrinkles, such as nasolabial folds and marionette lines or jowls.
Role of Lipids in the Human Body
Lipids can be found in different areas of the skin, not only in subcutaneous tissue. Lipids are constituents of phospholipids in the myelin sheaths of nerve tissue and cell membranes (lipid bilayers), play an important role in the skin barrier of the epidermis, and are essential for the production of steroids. They are water-insoluble organic molecules because they are nonpolar. However, after esterification (a condensation reaction between acid and alcohol), they are more water-soluble than their parent forms.
The most common lipids in the diet are triglycerides (triacylglycerol), which are composed of a glycerol subunit attached to three fatty acids (Fig. 3-3) Lipids can be saturated or unsaturated. Generally, an unsaturated fatty acid contains at least one double bond while saturated fatty acids do not. Unsaturated fatty acids provide slightly less energy during metabolism than saturated fatty acids with the same number of carbon atoms. In addition, saturated fatty acids are usually solids at room temperature
Apical layer
Dermis
Mantle layer
Deeper layer Muscle
Epidermis
FIGURE 3-1 The three layers of the subcutaneous tissue.
FIGURE 3-2 Android (apple) and gynoid (pear) fat distribution patterns in men and women.
Apple shape
Pear shape
and unsaturated fats are usually liquids at room temperature.
Lipid Metabolism
During digestion, fats in the food are broken down in the duodenum by pancreatic lipase into free fatty acids and glycerol. The intestinal epithelium absorbs these substances and reesterifies them in the smooth endoplasmic reticulum into triglycerides. These triglycerides are then absorbed into the circulation and lymphatic system. When they arrive in the circulation they are combined with apoprotein to form a lipoprotein, which is called a chylomicron. Chylomicrons are exposed to lipoprotein lipase, which is synthesized by adipocytes and stored at the surface of endothelial cells. Lipoprotein lipase cleaves the chylomicron into free fatty acids and glycerol again. These free fatty acids pass into adipocytes and combine with intracellular glycerol phosphate to form triglycerides and are stored for energy.
Adipose tissue can also convert excessive glucose and amino acids into fatty acids when stimulated by insulin. This explains why people who consume a low-fat diet or fat-free diet still gain weight if they do not reduce the total amount of calories they consume or have a high-carbohydrate diet. High blood glucose can stimulate insulin synthesis and insulin can increase synthesis of lipoprotein lipase from adipocytes to help absorb triglycerides into the cells. People who want to control their weight should avoid any foods that have the ability to stimulate insulin production. Individuals with type II diabetes have high levels of insulin; therefore, they have a higher risk of becoming overweight or obese than nondiabetic individuals.
Lipoproteins
There are many different types of lipoproteins. Low-density lipoprotein
(LDL) brings fat to the cells, while highdensity lipoprotein (HDL) brings fat from the circulation to the liver for excretion in bile. High levels of LDL are associated with a high incidence of coronary artery disease and atherosclerosis. HDL, or the “good lipoprotein,” can be elevated with exercise.
Lipid Synthesis
Triglycerides are derived from foods or synthesized from excessive glucose or amino acids. In humans, triglycerides are stored mainly in adipose tissue, which constitutes the body’s reserve energy source. However, excessive consumption of calories can lead to the synthesis and accumulation of more fat in subcutaneous tissues. Unfortunately, fat storage is unlimited in the subcutaneous tissue, unlike glycogen storage in the liver and muscle. Therefore, excessive fat accumulation will not only change a person’s cosmetic appearance but also increase their risk for osteoarthritis, diabetes, hypertension, as well as other diseases.
VOLUME EXCESS
Obesity
Obesity is defined as unhealthy, excessive fat mass. There are many regimens, products, and exercise programs available; however, there is still a rising pandemic in the United States9 when compared to the past.10 Obesity and hyperlipidemia are major risk factors and can lead to significant morbidity and mortality.
PATHOPHYSIOLOGY Obesity results from both environmental and genetic factors. Two genes that are known to have direct effects on obesity are the leptin ( ob gene )11,12 and proopiomelanocortin (POMC) genes.13 These genes can control eating behavior and satiety. Defects in these genes can cause severe obesity.
TABLE 3-1
Body Mass Index (BMI) Categories
BMI WEIGHT STATUS
Less than 18.5Underweight
18.5–24.9Normal
25.0–29.9Overweight
30.0 and greaterObese
However, almost all people gain weight when they get older because of diminished physical activity and aginginduced changes in the chemical activity of hormones.
Body mass index [BMI: body weight divided by the square of height (kg/m2)] is a popular index used for determining body weight status. The Centers for Disease Control and Prevention (CDC) and World Health Organization (WHO) use this index to classify adults into four groups (Table 3-1)
A normal BMI does not necessarily mean that a person has a “perfect” shape. Many people with a BMI less than 25 have fat accumulation in some area, such as the abdomen or buttocks.
IMPACT OF OBESITY ON THE SKIN
Obesity is responsible for changes in skin barrier function by significantly increasing transepidermal water loss, which can lead to dry skin and impaired barrier function.14 Hyperfunction of sebaceous glands due to high levels of androgenlike hormone or insulin-like growth factor hormone can aggravate severity of acne and hirsutism;15,16 delay wound healing and collagen deposits in the wound healing process;17 and disturb both blood and lymphatic circulation, which can cause angiopathy 18 and lymphedema, potentially precipitating chronic leg ulcers.19 Rapid weight gain can cause striae distensae (stretch marks), which are challenging to treat.20–23 In addition, in intertriginous areas such as the underarms, breasts, and groin, moisture accumulation can lead to candida infection (intertrigo).
It is widely known that obesity increases the risk of coronary heart disease, hypertension, hyperlipidemia, osteoarthritis, and diabetes. It is also known to be directly related to increased risk of sleep apnea; breast, endometrial, and colon cancer; gallbladder disease; musculoskeletal disorders; severe pancreatitis and diverticulitis; infertility;
FIGURE 3-3 Triglyceride chemical structure.
TABLE 3-2
Classification of Overweight and Obesity by BMI
BMI WEIGHT STATUS
25.0–29.9Overweight
30.0Obese
30.0–35.0Moderate obesity (Class I)
35.0–40.0Severe obesity (Class II)
40.0Morbid obesity (Class III)
urinary incontinence; and idiopathic intracranial hypertension. Additionally, obesity has indirectly been related to anxiety, impaired social interaction, and depression.
Obesity is implicated in a wide spectrum of dermatologic diseases, including acanthosis nigricans, acrochordons, keratosis pilaris, hyperandrogenism and hirsutism, striae distensae, adiposis dolorosa, fat redistribution, lymphedema, chronic venous insufficiency, plantar hyperkeratosis, cellulitis, skin infections, hidradenitis suppurativa, psoriasis, insulin resistance syndrome, and tophaceous gout.24
To determine the severity of a person’s obesity, BMI can be used. In fact, the more overweight a person is the higher the mortality rate (Table 3-2).
TREATMENT Dietary control is very important in the treatment of obesity. Patients must understand the principle of energy intake and expenditure. Weight reduction is usually not accomplished without exercise. However, exercise alone will usually produce little long-term benefit. The combination of exercise with dietary therapy can prevent weight being regained. In addition, regular exercise (30 min daily) will improve general health. The best results are obtained with education in well-motivated patients. Constant supervision by healthcare professionals and by family or friends can help to encourage compliance.
PREVENTION Prevention of obesity is key because once fat is gained and maintained over time, it is more difficult to lose. A high-fat diet can induce an increase in the number of adipocytes.25,26 A low-fat and complex carbohydrate diet is recommended to reduce body weight. There is an important difference between preventing weight gain and producing weight loss. To prevent weight gain, portion size and composition of food are controlled. For weight loss, restriction of calorie intake is the most effective treatment.
Liposuction
Overweight patients frequently consult plastic surgeons and dermatologists for liposuction.27–29 Liposuction is one of the most commonly performed cosmetic surgery procedures in the United States.30 Physicians must inform their patients that liposuction is a modality for improving body contour and not for treatment of generalized obesity. In addition, excess fatty tissue will return if regular exercise and diet control are not maintained.
Large-volume liposuction may decrease weight and fat mass; however, there is controversy regarding whether or not it significantly improves insulin resistance and other obesity-associated metabolic abnormalities.31–33 The most common areas treated are the neck, jowls, arms, abdomen, thighs, knees, and ankles. Other conditions that can be improved by liposuction include lipoma, gynecomastia, buffalo hump, and axillary hyperhidrosis. There are strict guidelines from both the American Society of Dermatologic Surgery (ASDS) and the American Academy of Cosmetic Surgery (AACS) on the volume restrictions during liposuction. Tumescent liposuction is considered the safest method for performing the pro-
cedure. This technique relies on the infiltration of dilute anesthesia based on body weight, and the removal of limited amounts of adipose tissue during each operation. Tumescent anesthesia consists of very dilute lidocaine and epinephrine solutions ranging from 0.05% to 0.1% of lidocaine with 1:1,000,000 epinephrine and sodium bicarbonate. The total safe concentration of lidocaine that can be used in this formula is 35 to 55 mg/kg based on patient weight and any coexisting medical conditions. Table 3-3 is a synopsis of the 2006 ASDS guidelines of care for tumescent liposuction.28
LIPOSUCTION COMPLICATIONS While there have been reports of mortality with general anesthesia, there have been no reports of death with tumescent anesthesia alone. When practitioners adhere to the AACS and ASDS guidelines, tumescent liposuction is a safe outpatient procedure. Common complications are bruising, swelling, localized paresthesia, and irritated incision sites after liposuction. Other complications include hematomas, seromas, and infection. There are serious complications that the surgeon must be aware of, however, such as the development of a fat embolus,
Indications
Aesthetic body contouring:most common regions include thighs,abdomen,hips,arms,back, buttocks,neck,breasts,and calfs
Other indications:treatment of lipomas,gynecomastia,lipodystrophy,axillary hyperhidrosis,axillary bromidrosis,and subcutaneous fat debulking during reconstructive procedures
Preoperative evaluation
History:diet patterns,exercise,unwanted regions,underlying disorders such as poor wound healing,bleeding abnormalities,diabetes mellitus,keloid formation,problems with past surgical procedures,personal or family history of thrombophlebitis,pulmonary emboli,and drugs that may interfere with blood coagulation or the metabolism of lidocaine
Explanation:procedure,risk and benefits,expected outcomes,needing a touch-up procedure
Physical examination:assessment of both general physical health and specific sites amenable to liposuction
Laboratory studies:may or may not be necessary for a given patient depending on the type and extent of anticipated liposuction procedure
Some surgeons may wish to obtain CBC,PT,PTT,LFT,UA,pregnancy test,screening for HIV, hepatitis B,and hepatitis C
Technique
Tumescent Anesthesia:consists of very dilute lidocaine and epinephrine solutions ranging from 0.05%–0.1% of lidocaine with epinephrine (around 1:1,000,000),sodium bicarbonate and / triamcinolone
Volume removal
Removal of more than 4 L of supranatant fat should be divided into more than one operative session
Monitoring:pulse oximetry,cardiac monitoring,and intermittent monitoring of BP,HR,and RR
Postoperative care
Use compression garments for 1 to 4 wk
TABLE 3-3
Synopsis of 2006 ASDS Guidelines of Care for Tumescent Liposuction