a primer of life histories : ecology, evolution , and application
A Primer of Life Histories
Ecology, Evolution, and Application
Jeffrey A. Hutchings
Professor of Biology, Department of Biology, Dalhousie University, Canada
1Great Clarendon Street, Oxford, OX2 6DP, United Kingdom
Oxford University Press is a department of the University of Oxford. It furthers the University’s objective of excellence in research, scholarship, and education by publishing worldwide. Oxford is a registered trade mark of Oxford University Press in the UK and in certain other countries
All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, without the prior permission in writing of Oxford University Press, or as expressly permitted by law, by licence or under terms agreed with the appropriate reprographics rights organization. Enquiries concerning reproduction outside the scope of the above should be sent to the Rights Department, Oxford University Press, at the address above
You must not circulate this work in any other form and you must impose this same condition on any acquirer
Published in the United States of America by Oxford University Press 198 Madison Avenue, New York, NY 10016, United States of America
British Library Cataloguing in Publication Data
Data available
Library of Congress Control Number: 2021934828
ISBN 978–0–19–883987–3 (hbk.)
ISBN 978–0–19–883988–0 (pbk.)
DOI: 10.1093/oso/9780198839873.001.0001
Printed in Great Britain by Bell & Bain Ltd., Glasgow
Links to third party websites are provided by Oxford in good faith and for information only. Oxford disclaims any responsibility for the materials contained in any third party website referenced in this work.
To my parents,Wendy and Alexander Hutchings
Preface
Science is a way of seeing the world through description, analysis, and interpretation of empirical patterns and processes. Many of the mechanics of science are technical in nature. One may need to know how to run a model simulation, isolate DNA, set a trap, raise seedlings, use a centrifuge, operate a boat engine. The technical demands of research are often obvious.
Less obvious is the need to identify a contextual, interpretative, and analytical framework that allows you to make sense of research findings and to draw conclusions about their potential utility or significance. Are there overarching principles, theories, or other generalities that would aid you in interpreting your research, communicating it to others, and increasing the probability that your work will in some small or large way advance knowledge and understanding? As a naïve master’s student in the early 1980s, I struggled to identify such a framework. I had little confidence in my ability to distinguish fundamentally important from fundamentally mundane questions.
My confidence received an unexpected boost by Stephen Stearns’ 1976 review on life-history evolution, written when he was a graduate student at University of British Columbia. It offered a fresh, taxonomically broad way of thinking about adaptation and natural selection. Why, indeed, should an organism reproduce once in its life and die immediately thereafter?
By 1992, sufficient life-history data were available that allowed for the testing of ideas, the poking of model assumptions, and the prodding of hypotheses. The stage was thus set for the first two general books on life-history evolution. Stearns wrote one; Derek Roff of McGill University wrote the other. Although bearing the same title (Evolution of life histories), there were differences in how topics were approached. Reaction norms figured prominently in Stearns’ contribution; quantitative genetics was emphasized by Roff. Joined by Roff’s Life history evolution (2002), these works contributed immeasurably to the torrid pace of life-history research that continues unabated (Figure 1).
Given this apparent enthusiasm, it seemed an opportune time to engage and hopefully enthuse new generations of students and researchers on the grandeur of life-history evolution, its theoretical underpinnings, and some practical applications. Comprising ten chapters, this primer is intended to be accessible to readers from a broad range of academic backgrounds and experience who have interests in ecology, evolution, conservation, or resource management.
Chapters 1 to 4 focus on core elements of life-history theory: population growth; trait variability; trade-offs; genetic architecture; reaction norms; reproductive effort; and reproductive costs. Chapter 5 offers tractable means of estimating fitness and predicting optimal changes in life history, using life tables. The next three chapters examine lifehistory evolution in variable environments, including bet-hedging (Chapter 6), theories
Figure P.1 Annual number of published papers whose titles, abstracts, or keywords included ‘life history’ or ‘life histories’ (hyphenated and non-hyphenated), according to the Web of Science, from 1900 to 2020.
for the evolution of offspring number and size (Chapter 7), and alternative reproductive tactics and strategies (Chapter 8).
Chapters 9 and 10 bridge the fundamentals of life-history theory to matters of applied interest from conservation and resource-management perspectives. Scaling up from individuals to species, Chapter 9 illustrates how life histories are inextricably linked to the vulnerability of species to extinction, exploitation, and climate change. Chapter 10 completes the primer with a look at how life histories affect sustainable rates of exploitation and how exploitation can, in turn, affect life histories.
The inspiration of this book is large enough. If it fails in its portrayal, the fault lies with an art that is deficient rather than an enthusiasm that is wanting.
Jeffrey A. Hutchings Halifax, Nova Scotia, Canada 8 January 2021 viii Preface
Acknowledgements
This primer had its genesis on a snowy winter evening in Skåne, Sweden, on 27 January 2014, the result of a lengthy conversation with Per Lundberg, professor of theoretical and evolutionary ecology at Lund University. Four years later, having decided I could create sufficient time to write, I travelled to Iceland to write chapter one in Auðunnarstofu, the outstanding fourteenth-century replica building serving as the office of my official host, the Bishop of Hólar, Solveig Lára Guðmundsdóttir. The trip, hosted also by my dear friend and colleague Skúli Skúlason (professor of fish biology at Hólar University), provided me with an ideal setting in which to think, walk, and write. Bjarni Kristófer Kristjánsson (professor and head of the department of aquaculture and fish biology, Hólar University) facilitated my interactions with the Icelandic academic and nonacademic communities, never failing to enlighten and inspire me with his cavernous knowledge of Icelandic history. In central Finland, where I wrote considerable portions of the book, I am indebted to Anna Kuparinen (professor of natural resources and environment, University of Jyväskylä) for her patience, support, intellect, and sagacity.
There are many others to whom I wish to offer my sincere thanks, admiration, and appreciation. First and foremost are those who reviewed portions, or the entirety, of one or more draft chapters: Hugues Benoît, Anna Kuparinen, Andrew Simons, Doug Swain, and Laura Weir. Several colleagues provided figures, photographs, data, published works, or unpublished manuscripts during the writing period, including Ken Andersen, Spencer Barrett, Eric Charnov, Larry Greenberg, David Hardie, Richard Law, Susan McRae, Julian Olden, Fanie Pelletier, Jeremy Prince, William Le Quesne, and Patrick White. I am very grateful to Jon Tremaine (Cornwall, UK) whose species-infused hare adorns the book cover. Ian Sherman and Charles Bath (Oxford University Press) faithfully and professionally advised me throughout the writing period.
I wish to acknowledge those who, perhaps unwittingly, provided mentorship and guidance as I strived to educate myself on matters pertaining to life-history evolution through the 1980s and 1990s: Graham Bell, Richard Law, Ransom Myers, Linda Partridge, Derek Roff, and Robert Wootton. Lastly, I owe my greatest thanks to my PhD supervisor Douglas Morris, not only for introducing me to Stearns’ (1976) review in 1983, but for impressing upon me the importance of intellectual honesty, academic integrity, respectful interactions with others, and asking questions of fundamental importance.
1.2
2.2.3
2.2.4
2.2.6
2.4.1
2.4.2
2.4.3
2.4.4
2.4.5
5.4
5.5
3.2.2
3.2.3
4.5.1
4.5.2
4.5.3
5.3.1
5.3.2
5.3.3
5.3.4
5.3.5
5.5.1
5.5.2
5.5.3
5.5.4
5.6
6.1 Shifts in the mean and variance of environmental conditions
6.2 Evolution of age and reproductive effort at maturity
6.2.1 Semelparity versus iteroparity
6.2.2 Life-history responses to changes in survival
6.2.3 Life-history responses to changes in the variance in survival
6.3 Life-history evolution in a variable environment
6.3.1 Hedging evolutionary bets
6.3.2 Reducing the variance in fitness
6.3.3 Bet-hedging life histories
6.3.4 Deterministic versus stochastic
6.4 Summing up and a look ahead
7 Number and Size of Offspring
7.1 Extreme variability in the production of propagules
7.2 Offspring number and size: not all options are possible
7.3 Evolution of offspring number
7.3.1 Early thinking: clutch size in birds
7.3.2 The Lack clutch size
7.3.3 Observed clutch sizes often differ from the Lack clutch size
7.4 Evolution of offspring size
7.4.1 Early thinking: a trade-off between size and number of offspring
7.4.2 Investment per offspring
7.4.3 Smith-Fretwell model of optimal size and number of offspring
7.4.4 Selection for increased fecundity
7.5 Summing up and a look ahead
8.1 Alternative reproductive phenotypes
8.2 Dichotomies in sex, size, and status
8.3 Thresholds and conditional tactics
8.4 Genetic polymorphisms
8.5 Genotype-by-environment thresholds for alternative strategies
8.5.1 Threshold reaction norms
8.5.2 Thresholds for alternative strategies under selection
8.6 Frequency-dependent selection
8.7 Clarity in terminology
8.8 Summing up and a look ahead
9 Applications: Conservation Biology
9.1 Three paradigms
9.2 A stimulus for incorporating life history to assess species vulnerability
9.3 Life-history correlates of r
9.4
9.6.3
9.7
10 Applications: Sustainable Exploitation of Evolving Resources
1 Fundamentals
1.1 A Brief History of Life Histories
1.1.1 Breadth
Two words are at the core of this book: ‘life’ and ‘history’. Considered singly, writing a primer on either would be presumptuous. But when combined, they offer the foundation of intellectually rewarding, scientifically tractable avenues of inquiry that are not specific to a particular taxon (plant or animal), biological scale (gene, genome, individual, population, species), or mechanism of change (physiology, development, plasticity, evolution).
To be intellectually engaged in the study of life histories from an ecological and evolutionary perspective is to be intellectually engaged in breadth. Not all scientists are comfortable with breadth. Some consider it unwise or unhelpful to stray from the comforts of a constrained set of theoretical and empirical constructs that anchor many research programmes. Others find breadth liberating, if not vital to maintaining their intellectual engagement in science over a 30- or 40-year career. If you are used to thinking broadly, tackling a narrow research question can be approached with confidence; if you are used to thinking narrowly, tackling a research question of breadth can be daunting.
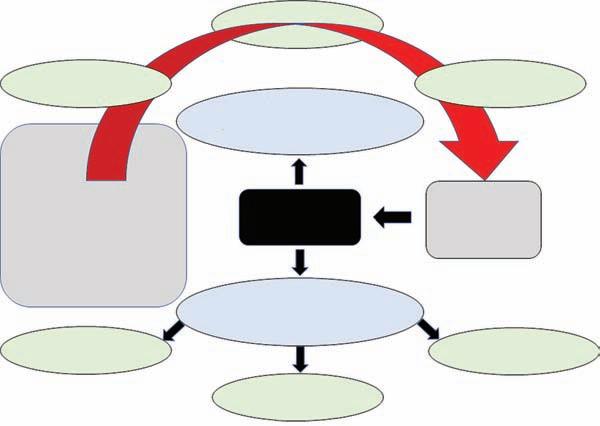
To get a feel of the inherent breadth of life-history research, consider a simple figure that illustrates how various factors influence individual life histories, along with some of the consequences these can have at the population level (Figure 1.1). Rather than a mechanistically or formatively accurate flow diagram, think of this figure as a roughly organized cork board into which various constituent elements of life histories have been pinned. Note that none of the elements is specific to a particular taxonomic group; this permits breadth of interest and breadth of inquiry. The box on the left subsamples a range of traits and processes that comprise an individual’s phenotype, i.e. its observable form. Following the curved arrow, we are reminded that these traits and processes are influenced, through selection, by an individual’s genes, the environment it experiences, and interactions between genotypes and their environment.
The end result is a combination of life-history traits that determines an individual’s probability of surviving to, and reproducing at, various ages or stages—its life history. As
Figure 1.1 Diagrammatic representation of factors that influence life history which ultimately has direct implications for individual fitness. Correspondingly, individual fitness affects individual and population rates of increase which have consequences for applied issues related to sustainable exploitation and conservation.
we shall see in this chapter, an individual’s life history determines individual fitness (the rate of increase in that individual’s genes through time). From an applied and conservation perspective, the average fitness of individuals in a population can affect things such as sustainable rates of exploitation (fishing, forestry, hunting), speed and uncertainty of species/population recovery following depletion, and the ability of a population to persist following natural or human-induced disturbance.
1.1.2 ‘Life history’ in the nineteenth century
The concept of a life history today differs from that of the 1800s when a life history was equivalent to a life cycle. Embryo to egg to larva to adult. Seed to seedling to later vegetative, flowering, and pollination stages. A life history was a description of the development of the presumed generic ‘type’ of a particular species.
That said, scientific thinking of life histories in the nineteenth century was not entirely devoted to descriptive summaries alone. Charles Darwin (1859) and Ernst Haeckel (1866) were among those who thought evolution to be involved insofar as it could affect things such as the length and number of life stages and their associated morphological and developmental features. After recognition of Georg Mendel’s work on plant hybridization (1856–1863) at the turn of the twentieth century, scientists began to explicitly link life histories to genetics. As one example, in 1909 Adam Sedgwick (whose greatuncle of the same name guided Darwin’s early studies) asked:
What is the relation of these [developmental] variations in structure, which successively appear in an organism and constitute its life-history, to the mutational variations which appear in different organisms of the same brood or species? (Sedgwick 1909: 181)
As the 1800s eased into the 1900s, life-history studies were very much focused on individual species. There was a paucity of ecology, evolution, and genetics. There was a paucity of breadth. Things were about to change.
1.1.3 The turning point: 1930
Ronald Fisher was pivotal in the development of modern statistical science and mathematical population genetics. Less well-appreciated are his foundational contributions to research on life-history evolution. In the opening chapter of The genetical theory of natural selection, he argued that researchers who have accepted the concept of natural selection will ‘direct [their] inquiries confidently towards a study of the selective agencies at work throughout the life history’ of organisms (Fisher 1930: 21). It marked a pivotal turning point in the development and application of life-history theory.
Fisher bridged the gap between the nineteenth- century concept of a life history and the early twentieth-century concept of natural selection. To do so, he seized a tool long- used in studies of human population growth: ‘To obtain a distinct idea of the application of Natural Selection to all stages in the life- history of an organism, use may be made of the ideas developed in the actuarial study of human mortality’ (Fisher 1930: 22).
The ‘ideas’ Fisher was referring to were human actuarial tables. Originally developed in the early 1800s for life-insurance purposes, these tables summarized the probabilities that humans survive from one age to subsequent ages. Fisher went further, arguing that a life table of survival was, in itself, ‘inadequate to express fully the relation between an organism and its environment; it concerns itself only with the chances of frequency of death, and not at all with reproduction’ (Fisher 1930: 24). By combining probabilities of age-specific survival with age-specific reproduction, he produced what ecologists and population biologists today would term a ‘life table’. Fisher showed how one could readily calculate the number of offspring that each newly born individual, or more precisely ‘genotype’, would be expected to produce over that individual’s lifetime.
So, in addition to accounting for the probability of surviving from birth to subsequent ages, Fisher accounted for the rate of reproduction at each age. If each individual in a population produced, on average, sufficient numbers of offspring to exactly replace themselves, the population would remain stable. However, if the per individual or ‘per capita’ production of offspring surviving to maturity was greater or less than one (i.e. replacement), the population would increase or decline, respectively. Fisher defined the per capita production of offspring as the Malthusian parameter of increase (named after Thomas Malthus whose An essay on the principle of population (1798) heavily influenced Darwin and Alfred Wallace).
Fisher argued that the per capita rate of increase should be directly linked to the strength of natural selection. The greater the per capita production of offspring by individuals of
A Primer of Life Histories
a particular genotype, relative to the rate of other members in the same population, the greater the strength of selection.
The necessity of accounting for reproduction when exploring how natural selection acts raised questions concerning the effort an individual should expend on reproduction. What are the consequences to future survival and reproduction associated with the present allocation of greater or lesser amounts of reproductive effort? An oft-quoted sentence from Fisher draws the reader’s attention to these potential trade-offs:
It would be instructive to know not only by what physiological mechanism a just apportionment is made between the nutriment devoted to the gonads and that devoted to the rest of the parental organism, but also what circumstances in the life-history and environment would render profitable the diversion of a greater or lesser share of the available resources towards reproduction. (Fisher 1930: 43–4)
Assuming that the resources available to an individual at any given age are fixed, their diversion to some facet of reproduction must come at the expense of not allocating those same resources to components of survival and future reproductive capacity, such as body maintenance, growth, metabolism, foraging, and vigilance. In other words, present reproductive effort likely comes at a future reproductive cost. Fisher speaks directly to his underlying postulate that the allocation of resources to reproduction, as opposed to other things, is related to ‘circumstances in the life-history and environment’, i.e. natural selection.
By linking the nineteenth-century, evolutionarily mute, stage-based concept of a life history to his twentieth-century ideas of how natural selection acts (through genetic processes) on age-based probabilities of survival, Fisher brought logical and mathematical clarity to how the strength of selection acting on genotypes varies with age and developmental stage.
A new definition of what constitutes a life history emerged from his work, one that can be expressed in the following way: A life history describes how genotypes vary their age- or stage-specific expenditure of reproductive effort in response to extrinsic and intrinsic factors that affect age-specific survival and fecundity. Or, more succinctly, life histories are the probabilities of survival and the rates of reproduction at each age in a lifespan (Partridge and Harvey 1988).
1.1.4 Fifty years on (1930–1980)
Fisher offered a turning point from nineteenth- to twentieth-century thinking about lifehistory variability and how natural selection acts on this variation. He laid the foundation for life-history theory. But it was left to others to construct the explanatory and predictive frameworks for understanding why individuals differ so extraordinarily, within and among species, in the means by which they propagate genes to future generations. These efforts were stimulated by fundamental questions of life-history evolution, examples of which are provided as points of departure for cameos that illustrate key developments in the fifty years that elapsed since the publication of Fisher’s book.
Question: Why do some organisms breed multiple times, whereas others breed once and then die?
Many organisms reproduce iteratively, or multiple times, throughout their lives. Others breed once and die shortly thereafter. LaMont Cole (1954), who termed these lifehistory patterns iteroparity and semelparity, respectively, was the first to explore their fitness consequences.
Although Cole (1954) examined several facets of life histories, researchers have been somewhat obsessed with his ‘paradox’ (sub-section 6.2.1). Surprisingly, he concluded that a semelparous population that produced one offspring more than the mean fecundity or number of offspring (b) of an iteroparous population would have the same rate of increase as the iteroparous population. (Put another way, a semelparous population with mean fecundity of (b + 1) would have the same rate of increase as an iteroparous population with mean fecundity b.) While adding a single offspring to a bird’s clutch of five might sound plausible in rendering the fitness of semelparity equal to that of iteroparity, the same cannot be said for adding a single offspring to the million produced by a fish. The paradox is that if an extremely small increase in fecundity should favour semelparity over iteroparity, why is semelparity so uncommon? It turns out that Cole’s model incorporated the assumption that iteroparous individuals experience no mortality, not even as very small eggs or seeds. Cole’s paradox was solved when empirically realistic rates of juvenile mortality were incorporated and the costs of semelparity were rendered more intuitively reasonable (Charnov and Schaffer 1973; Bell 1976).
Lamentably, Cole’s paradox has had a tendency to overshadow his other formative contributions to life-history evolution. A mathematically skilled ecologist (he worked on reptiles), Cole was the first researcher to have overtly concurred with Fisher’s perspective on the evolution of life histories, agreeing that ‘any life history features affecting reproductive potential are subject to natural selection’ (Cole 1954: 104). His classic 1954 paper—The population consequences of life history phenomena—was the first since Fisher’s book to include the words ‘life history’ in the title; among the ~310 papers that cited Fisher (1930) between 1930 and 1955, Cole’s is the only article to do so (according to Google Scholar).
Question: Why do some organisms start breeding early in life and others comparatively late in life?
The key to answering this question is to understand how age at maturity affects fitness. Cole (1954) explored this issue in detail, demonstrating mathematically that fitness is highly sensitive to the age at which an organism first reproduces, especially at comparatively young ages. All else being equal, the younger you start reproducing, the higher your fitness. But, because of life-history trade-offs, such as the allocation trade-off mentioned in sub-section 1.1.3 (also, see Chapter 4), all else is rarely equal when it comes to comparing the fitness associated with alternative ages at maturity.
Despite the existence of trade-offs, age at maturity retains a dominant influence on fitness. Garth Murphy (1968), who used models of Atlantic herring (Clupea harengus) to poke and prod Cole’s paradox, concluded that age at maturity depends on the ratio, and the stability, of survival during the juvenile (pre-reproductive) stage relative to survival during the adult (reproductive) stage (sub-sections 5.5.4 and 6.2.2). Murphy’s work
A
Primer of Life Histories
provided the basis for predicting how changes in survival and fecundity affect the evolution of age at maturity.
Age at maturity is a life-history trait. Cole (1954) may have been the first to explicitly articulate this connection, referring to it as a life-history ‘feature.’ He also identified fecundity and longevity as life-history features/traits and predicted how they might affect rates of population change. At about the same time, some empirically-minded ecologists were honing in on another life-history trait—offspring size.
Question: Why do some organisms produce many, small offspring and others few, large offspring?
For a fixed amount of ‘effort’ there is a clear trade-off between the number of offspring produced and the size of each of those offspring, be they seeds, eggs, or embryos. If you are interested in explaining the adaptive significance of variability in one of these two traits (offspring size and offspring number), you will find yourself on intellectually thin ice if you do not account for variability in the other (more of that in Chapter 7).
Early attempts to understand inter-specific variability in offspring number within an evolutionary context can be traced to the ornithologist David Lack whose work in the late 1940s centred on altricial birds (young fed by their parents at a nest). By assuming that the food that parents can feed their young is limited, Lack (1947a) reasoned that the number of young produced cannot increase without a reduction in the amount of food provided to each of those young. In other words, the number of eggs laid reflects the optimal number of young that the parents can feed and/or provide parental care for.
At the same time Lack was pondering the evolution of clutch size in birds, Gunnar Svärdson, a Swedish fish biologist, was considering the evolutionary implications of how offspring size affects offspring number. Foreshadowing what was to come in the early 1970s, he suggested there must be an upper limit to offspring number that depended on how offspring size was related to offspring survival and parental reproductive success (Svärdson 1949). Twenty-five years later, Christopher Smith and Stephen Fretwell (1974), using graphical models, provided an analytical solution for determining the optimal balance between offspring size and offspring number (sub-section 7.4.3). Their simple model remains the starting point today for most explorations of the evolution of egg/seed size.
Question: How costly is reproduction?
The logical necessity of reproductive costs is based on the premise that organisms are energetically constrained systems. If energy required for growth, maintenance, and reproduction originates from the same fixed pool of resources, an allocation of energy to reproduction cannot be made without a reduced allocation to other body functions or activities (Chapter 4).
The genetic basis for reproductive costs was initially explored by Peter Medawar (1952) and George Williams (1957). In discussing the evolution of ageing, or senescence, they argued that natural selection may favour a gene that has beneficial effects early in life even if the same gene greatly reduces fitness later in life. Williams (1966) extended this theory to reproductive effort, reasoning that if energy allocation to present reproduction reduces future reproductive success, this cost of reproduction will result in
the evolution of life histories that are characterized by intermediate levels of survival and reproduction.
So, the greater the effort, the greater the subsequent cost. While such a causal link seemed self-evident, it was empirically unclear what this relationship might actually look like. The shape of the function relating reproductive effort to reproductive cost can strongly influence life-history evolution. This is implicit in all theories that are based on the existence of reproductive costs. For example, different shapes of cost functions can lead to the evolution of either iteroparity or semelparity or some combination of the two (Gadgil and Bossert 1970; Schaffer and Rosenzweig 1977; Bell 1980). Although cost functions are central to life-history theory, few have been empirically described. Measuring the magnitude of reproductive costs remains a significant empirical challenge.
1.1.5 Life-history ‘classics’
The preceding sub-sections offered a brief history of the core elements of life-history theory to 1980, by which time the foundations of modern approaches had essentially been set. Brevity inevitably involves exclusion and some life-history ‘classics’ were not mentioned. The interested student and researcher might wish to delve deeper into these foundational papers and a small sample of other fundamentally instructive texts published after 1980 (Table 1.1). (Be it art, music, literature, or science, what constitutes a classic is in the eye of the beholder.)
Table 1.1 Life-history topics and suggested foundational literature sources.
Topic Foundational papers or books
Semelparity vs iteroparity
Cole (1954); Murphy (1968); Gadgil and Bossert (1970); Charnov and Schaffer (1973); Schaffer (1974a, b); Bell (1976); Schaffer and Rosenzweig (1977); Young (1981)
Age at maturity Alm (1959); Hamilton (1966); Murphy (1968); Stearns (1976); Charlesworth (1980); Roff (1984); Stearns and Koella (1986)
Reproductive effort Tinkle (1969); Hirshfield and Tinkle (1975); Pianka and Parker (1975); Charlesworth and León (1976); Schaffer and Rosenzweig (1977); Goodman (1984); Charnov et al. (2007)
Offspring size and number Lack (1947a, b); Svärdson (1949); Cody (1966); Harper (1967); Janzen (1969); Harper et al. (1970); Smith and Fretwell (1974); Brockelman (1975); Capinera (1979)
Costs of reproduction
Medawar (1952); Williams (1957, 1966); Law (1979); Michod (1979); Bell (1980); Rose and Charlesworth (1981); Reznick (1985)
General Fisher (1930); Lewontin (1965); Williams (1966); Stearns (1976, 1992); Charlesworth (1980); Partridge and Sibly (1991); Roff (1992, 2002)
1.2 A Primer of Population Growth
1.2.1 Intrinsic rate of natural increase
Fisher (1930) placed mathematical population biology firmly at the forefront of scientific research on life-history evolution. Yet, as vital as mathematics has been to the development of life-history theory, the tendency for many biologists—young and old—to eschew quantitative analytics might well have served (and continue to serve) as a retarding force in life-history research, particularly in the empirical testing of life-history theory and its practical applications, of which there are multiple examples (Chapters 9, 10).
As early as the mid-1950s, Cole (1954: 135) was lamenting that studies of life histories have been ‘neglected from the evolutionary point of view, apparently because the adaptive values of life-history differences are almost entirely quantitative’. Derek Roff (1992: 3) concluded much the same thing almost 40 years later: ‘An early antipathy to the use of mathematical analysis may account in part for the delay in the merging of the ecological and evolutionary perspectives in what is now commonly known as “life history analysis” ’.
Fisher clearly did not share this antipathy, embracing Alfred Lotka’s (1907; Sharpe and Lotka 1911) model for continuous population growth and applying it within a lifehistory context. In a closed population (no immigration or emigration) growing at discrete time intervals (all births and deaths occurring at the same time every year), the number of individuals at time step t + 1 (Nt+1) can be expressed as the number of individuals at time t (Nt) plus the number of individuals born at time t (Birthst) minus the number of individuals that died at time t (Deathst): +
When modelling the change in population size from time step t to time step t+1, it is generally assumed that the likelihood of an individual giving birth or dying in that time interval will be fairly constant. These are termed the per capita rates of birth (b) and death (d), such that Births = bN (a rearrangement gives b = Births/N) and that Deaths = dN (and d = Deaths/N). Substituting these per capita rates into a slightly rearranged Equation 1.1 yields the following expression:
1.2
which can be rewritten as
The parenthetical term in Equation 1.3 is a constant multiplier of population change. It is usually termed the discrete or finite rate of population growth, simplified as λ, such that:
Equation 1.4
Thus, λ is the proportional rate of change in population size from one discrete time step to the next discrete time step, such that:
Although the use of discrete time intervals can be empirically defended, insofar as germination/breeding often occurs at a specific time each year for many species, deaths need not be similarly timed, often occurring continuously.
Mathematically, it can be convenient to simplify population growth as a process that occurs continuously, meaning that changes in population size can be modelled as occurring over extremely small intervals of time known as ‘instantaneous change’. A change in population size (N) over such an infinitesimally short period of time (∂t) is represented by ∂N/∂t.
In continuous time, as described above, the number of births is a function of population size, the instantaneous per capita birth rate (b = Births/N), and the instantaneous per capita death rate (d = Deaths/N). Thus, in continuous time, population growth rate is described by:
Note that this instantaneous rate of population change is controlled by the difference between the per capita rates of birth and death. Lotka (1907: 22) defined this difference as:
terming r the ‘rate of natural increase per head’ or the per capita rate of natural increase. Substituting Equation 1.8 into Equation 1.7 yields the standard model for population growth rate in continuous time:
which means that
1.10
Fisher (1930) defined r similarly (as have most life-history researchers since) but he called it the Malthusian parameter of population increase. According to Cole (1954), Lotka was inconsistent in what he called r, variously describing it as the ‘true’, ‘incipient’, ‘inherent’, and ‘intrinsic’ rate of increase. Cole, among others (e.g. Birch 1948), settled on ‘intrinsic rate of natural increase’. This is generally what r has been called since.
The ecological literature on λ and r can sometimes be confusing because the parameters often seem to be used interchangeably. But they do differ in important ways. Lambda (λ) is measure of population growth rate, i.e. the change in population size from one time step to another, whereas r is a measure of per capita population growth rate. Lambda can be thought of as the average contribution of each individual alive at time t to the size of the population at time t + 1, whereas r is the average contribution of each individual to the rate of change in population size. The two parameters are related to one another: λ = er and r = ln(λ).
1.2.2 Density-independent population growth
Whatever one calls r, it is quite important to remember that it is the per capita population growth rate. Consulting Equation 1.10, the units of r are ‘individuals per individual per unit of time’. By contrast, population growth rate (Equation 1.9), represented in continuous time by ∂N/∂t, has units of ‘individuals per unit of time’. Failure to distinguish per capita population growth from population growth is inexcusably common in the scientific literature.
The discrete (Equation 1.5) and continuous (Equation 1.9) models of population growth are both geometric or exponential functions. We describe this exponential growth by an equation that plots changes in population size over time, such that:
Equation 1.11
where N0 is the starting population size. Under these circumstances, we find N increasing steadily without bounds (Figure 1.2). This pattern of change in N with t is also called density-independent population growth.
Under density-independent population growth (Figure 1.2), the per capita population growth rate, r, remains constant; it does not change with population size (N). Rather, it remains at a maximal level, rmax, the maximum per capita population growth rate (dashed line in Figure 1.3). To improve clarity, Equation 1.9 can be better expressed as:
Equation 1.12
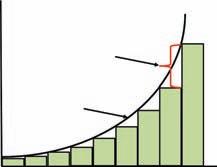
Figure 1.2 Exponential population growth over discrete time intervals (boxes, illustrating population change once per time step, t) and over continuous time (curved line) from an initial population size of N0.
Figure 1.3 For the density-independent model of population growth, r does not change with population size, N. It remains constant at the maximum value, rmax.
Thus, under exponential growth, the population size at any time t (Nt) depends on two parameters (numbers that remain constant) and one variable (numbers that take on different values), such that Nt depends on the starting population size (N0, a parameter), the maximum intrinsic rate of increase (rmax, also a parameter), and time (t, a variable):
Equation 1.13 (analogous to Equation 1.11) produces a curve similar in shape to that in Figure 1.2. This basic equation is used to model exponential changes in population size with changes in time. (The phrase exp(rmaxt) has come to mean to exponentiate the bracketed term, such that exp(x) = ex.)
1.2.3 Density-dependent population growth
Under the density-independent model (exponential growth), birth and death rates are assumed to be constant. Indeed, there is good empirical evidence that populations can experience exponential growth when their size is small relative to the numbers of individuals that their environment can sustain, i.e. their carrying capacity (K ).
As closed populations increase in abundance or density, their per capita growth rate, r, inevitably changes as competition for increasingly limiting resources, such as space and food, becomes increasingly intense. Now it becomes useful to distinguish ‘realized’ per capita growth (rrealized) from maximum per capita growth (rmax). Increased competition can have the effect of reducing the realized per capita birth rate and/or increasing the realized rate of per capita death. This means that as N increases, rrealized (which equals b – d), must decline (Figure 1.4).
The simplest means of incorporating an effect of increasing density (higher N in a closed population) on population growth rate is to reduce ∂N/∂t by an amount proportional to the remaining ‘portion’ of the carrying capacity (K), such that: